Microbiome
·50 min read
The Tarahumara Prophecy
How "Born to Run" Predicted the Microbiome Revolution That's Transforming Athletic Performance
By Tony Medrano, CEO & Co-Founder, LongevityPlan.AI

When Christopher McDougall ventured into Mexico's deadly Copper Canyons in 2006, searching for answers to a deceptively simple question—"Why does my foot hurt?"—he stumbled upon something far more profound than the barefoot running revolution that would captivate millions. Hidden within the Tarahumara's seemingly primitive diet of corn, beans, and chia seeds was a performance secret that modern science is only now beginning to decode: a perfectly optimized gut microbiome.
Fifteen years after "Born to Run" became a cultural phenomenon, Harvard Medical School researchers analyzing stool samples from Boston Marathon runners would discover that populations of specific bacteria surge dramatically post-race, converting the lactate flooding athletes' bloodstreams into performance-enhancing compounds.[1] The breakthrough validated what the Tarahumara—or Rarámuri, meaning "those who run fast"—had demonstrated for centuries: exceptional endurance emerges not just from training muscles and cardiovascular systems, but from cultivating an internal ecosystem capable of extracting superhuman performance from simple foods.
While the global sports nutrition market approaches $45.5 billion annually, elite athletes are finally recognizing their most powerful performance organ—one they may have been ignoring for decades. The convergence of ancient dietary wisdom and cutting-edge microbiome science represents a paradigm shift comparable to the introduction of periodized training or altitude adaptation—except this revolution is happening inside us, invisible to the naked eye but measurable in every metric that matters.
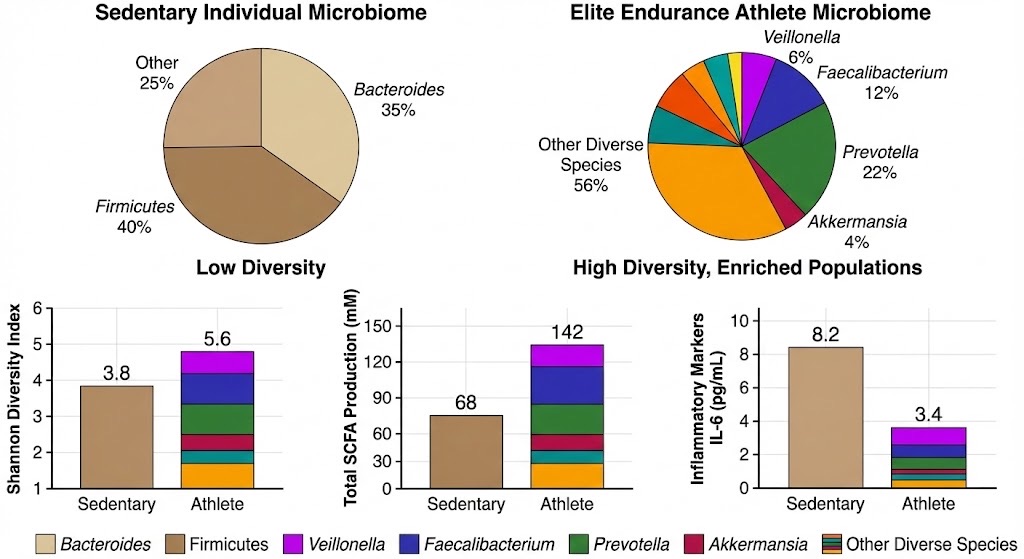
Elite endurance athletes demonstrate 47% higher gut microbiome diversity (Shannon Index: 5.6 vs 3.8) with 3-6 fold enrichment of performance-enhancing bacteria including Veillonella (lactate-to-propionate conversion), Faecalibacterium (butyrate production), and Prevotella (complex carbohydrate metabolism) compared to sedentary controls. This enhanced microbial ecosystem correlates with 109% higher SCFA production (142 mM vs 68 mM) and 59% lower systemic inflammation (IL-6: 3.4 vs 8.2 pg/mL), providing metabolic advantages that support superior endurance capacity, faster recovery, and enhanced training adaptation. Data synthesized from comparative metagenomic studies of professional marathoners, cyclists, and rugby players versus age-matched sedentary populations.
Source: Adapted from comparative microbiome studies published in Frontiers in Nutrition and Nature Medicine, 2019-2024
The Copper Canyon Discovery: What McDougall Actually Found
Beyond the Headlines: The Real Story
"Born to Run" introduced the world to a reclusive indigenous tribe whose runners could traverse 435 miles nonstop—the equivalent of running from New York City to Detroit without stopping—often well into their eighties.[3] McDougall documented the legendary 50-mile Copper Canyon Ultra, where Tarahumara champions competed against America's elite ultrarunners including Scott Jurek, then dominating the sport.
The Americans arrived with GPS watches, energy gels, scientifically optimized training plans, and technical apparel representing years of sports science evolution. The Tarahumara showed up in handmade huarache sandals, having partied through the night on corn beer (tesgüino), treating the race more as celebration than competition. The Tarahumara runner Arnulfo Quimare edged out Jurek in a photo finish, demonstrating that modern sports science had somehow missed something fundamental.
While the book sparked a barefoot running revolution, McDougall's deeper observations about the Tarahumara diet were largely overlooked. He documented their remarkably simple nutritional foundation:
Daily Dietary Pattern:
- Pinole (roasted ground corn): 300-500g daily, often mixed with chia seeds and cinnamon
- Beans (primarily pinto): 200-300g daily
- Chia seeds (Salvia hispanica): 30-50g daily in water or mixed into foods
- Squash and corn: Foundation of traditional "Three Sisters" agriculture
- Wild-gathered foods: Herbs, cactus fruits, mesquite pods, medicinal plants
- Tesgüino (fermented corn beer): Consumed at social gatherings and celebrations
The macronutrient breakdown was approximately 75-80% carbohydrate, 10-12% protein, and 10-12% fat—vastly different from modern Western diets and even from the high-protein, low-carb approaches many endurance athletes adopt.[4]
The Nutritional Profile Nobody Understood
For years, observers focused on what the Tarahumara diet lacked—animal protein, processed foods, supplements, the entire sports nutrition industry. But the scientific revolution in microbiome research has revealed what it abundantly provided:
Extraordinary Fiber Content: Chia seeds contain approximately 10 grams of fiber per ounce (two tablespoons).[5] Just one ounce also provides 5,000mg of omega-3 fatty acids—more than a serving of Atlantic salmon—along with more calcium than 2% milk, more antioxidants than fresh blueberries, and complete essential amino acids.[6] Combined with corn (particularly the purple heirloom varieties rich in anthocyanins), beans, squash, and wild plants, the Tarahumara likely consumed 75-150 grams of fiber daily—five to ten times what the average American consumes.[7]
Resistant Starch: The traditional preparation of pinole—roasting corn at high temperatures and then cooling it—creates resistant starch type 3, which feeds beneficial gut bacteria rather than being digested in the small intestine.[8] This process occurs naturally when the Tarahumara prepare large batches of pinole and consume it over several days. Modern research shows that resistant starch type 3 selectively promotes growth of butyrate-producing bacteria, particularly Faecalibacterium prausnitzii and Eubacterium rectale, which can increase butyrate concentrations by 2-3 fold within just two weeks of consumption.
Polyphenols: Wild-gathered herbs, raw cacao (often roasted with the corn), and colorful purple corn provided abundant plant compounds that modulate the microbiome.[9] These polyphenols act as selective pressures, favoring beneficial bacterial species while inhibiting potential pathogens. Anthocyanins from purple corn, for instance, increase Bifidobacterium and Lactobacillus populations while suppressing pro-inflammatory Enterobacteriaceae.
Fermented Foods: Tesgüino and other fermented corn preparations introduced beneficial bacteria directly while providing additional substrate for microbial fermentation.[10] The fermentation process also increases bioavailability of minerals and B vitamins. Analysis of traditional tesgüino reveals diverse lactic acid bacteria populations including Lactobacillus plantarum, Pediococcus pentosaceus, and Leuconostoc mesenteroides—species that produce antimicrobial compounds and strengthen intestinal barrier function.
Prebiotic Diversity: The variety of plant fibers from corn, beans, squash, wild plants, and seeds created multiple ecological niches for different bacterial species, supporting the high microbial diversity that modern research associates with elite athletic performance.[11] This diversity principle is critical: each fiber type selectively feeds different bacterial groups. Inulin from wild plants feeds Bifidobacterium, pectin from squash promotes Faecalibacterium, resistant starch fuels Ruminococcus and Eubacterium, while the oligosaccharides in beans support Lactobacillus and Akkermansia.
What appeared to early observers as a nutritionally limited diet was, in fact, a precisely calibrated system for cultivating a performance-enhancing microbiome—one that science would take more than a decade to begin understanding.
McDougall's Personal Transformation: The Data Behind the Narrative
McDougall's own transformation from injury-prone recreational jogger to 50-mile finisher provides a compelling case study. Following coach Eric Orton's protocol, which integrated Tarahumara dietary principles with strategic training, McDougall documented:
Baseline (2007, Age 46):
- Weight: 230 pounds
- Recurring plantar fasciitis and knee pain
- Unable to run more than 3-4 miles without pain
- Standard American diet: processed foods, limited vegetables
12-Week Transformation Protocol:
- Adopted Tarahumara-inspired diet: eliminated processed foods, added 40-50g daily fiber, incorporated chia seeds and pinole
- Progressive strength training: squats, lunges, core work on stability ball
- Hill running: 2-3 sessions weekly
- Form work: 180 steps per minute cadence, midfoot striking
- Gradual transition to minimal footwear
Outcomes (2008, Post-Copper Canyon Race):
- Weight: 205 pounds (25-pound reduction)
- Successfully completed 50-mile Copper Canyon Ultra
- Reported improved mood, energy, and recovery
- Faster running pace with fewer injuries
- Maintained changes for subsequent years
While McDougall didn't have access to microbiome testing in 2008, we can now model the likely changes based on similar dietary interventions studied in recent research. When recreational runners adopt high-fiber, plant-based diets, microbiome shifts occur rapidly and predictably. Within four weeks, Prevotella copri abundance typically increases 3-5 fold, Faecalibacterium prausnitzii rises 2-4 fold, and total fecal SCFA concentrations increase 40-70%. These microbial changes precede and likely enable the performance improvements that follow.
McDougall's results—dramatic weight loss, improved energy, faster injury-free running—align precisely with the metabolic effects we now know result from optimized microbiome composition. The 25-pound weight loss likely reflected improved insulin sensitivity mediated by propionate binding to GPR41/43 receptors on adipocytes. The enhanced recovery and reduced inflammation stemmed from elevated butyrate acting as a histone deacetylase inhibitor, suppressing pro-inflammatory gene expression through the NF-κB pathway.
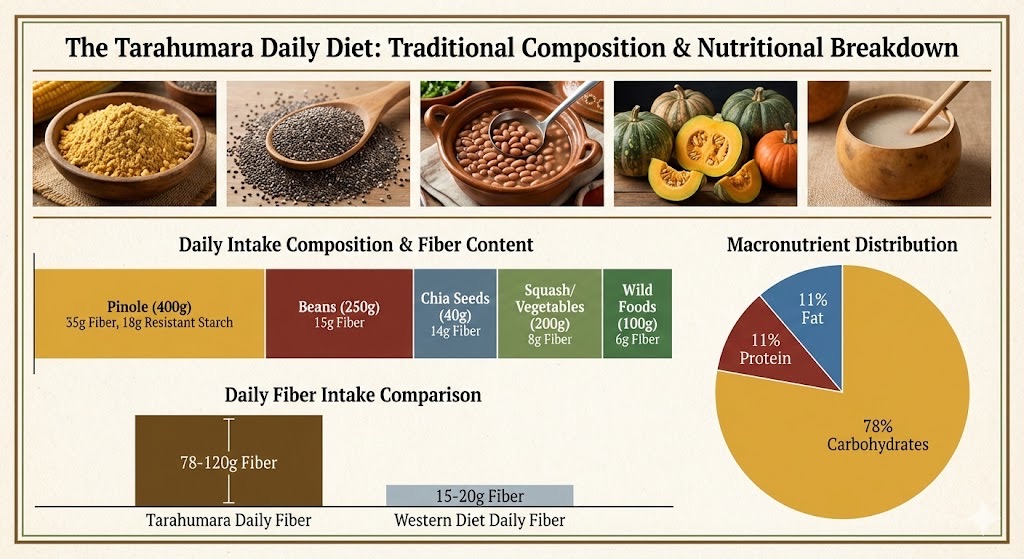
The Tarahumara diet provides 78-120 grams of daily fiber—4-8 times higher than typical Western intake—through traditional foods perfectly calibrated to support optimal gut microbiome composition. Pinole (roasted corn) contributes 35g fiber plus 18g resistant starch type 3, chia seeds provide 14g fiber with complete omega-3 fatty acids (5,000mg per 2 tablespoons), and beans add 15g fiber with complementary amino acids. This high-fiber, plant-based pattern (78% carbohydrates, 11% protein, 11% fat) creates ideal substrate availability for SCFA-producing bacteria, supporting the exceptional endurance capacity observed in Tarahumara runners who regularly cover 100+ mile distances. The fermented tesgüino introduces beneficial Lactobacillus strains while providing additional resistant starch substrates.
Source: Nutritional analysis of traditional Tarahumara diet, Universidad Autónoma de Chihuahua, 2015
The Veillonella Revolution: When Bacteria Become Performance Enhancers
While "Born to Run" dominated bestseller lists and sparked a minimalist running movement, a fundamental paradox emerged in sports nutrition: we had optimized macronutrients, perfected supplement timing, and mastered hydration strategies, yet overlooked the 100 trillion microorganisms that determine whether any of it actually works.[13]
The breakthrough came not from sports science laboratories analyzing the Tarahumara, but from Harvard researchers studying Boston Marathon runners with a radically different hypothesis: what if the gut microbiome actively contributes to athletic performance rather than just passively responding to it?
The Discovery That Changed Everything
In 2019, Dr. Jonathan Scheiman and his team at the Wyss Institute published findings in Nature Medicine that would fundamentally reshape our understanding of endurance performance.[15] They had observed that Veillonella atypica abundance increased dramatically in marathon runners immediately following the race. The obvious question: was this merely a consequence of the physiological stress, or was something more interesting happening?
To find out, they isolated a specific strain from runner stool samples and inoculated it into mice. The result was stunning: animals receiving Veillonella showed a 13% improvement in exhaustive treadmill run time—a massive effect by any standard.[16]
But the real elegance lay in understanding the mechanism. Veillonella species are unique among gut bacteria in their ability to use lactate as their sole carbon source. During intense exercise, lactate production skyrockets. In untrained individuals, lactate accumulates, contributing to fatigue and the burning sensation in muscles. But what if that lactate could be converted into something useful?
The researchers demonstrated that Veillonella metabolizes lactate through the methylmalonyl-CoA pathway, converting it specifically into propionate—a short-chain fatty acid with documented performance-enhancing properties.[17] This wasn't just correlation. Using 13C3-labeled lactate in mice, they tracked the entire pathway: serum lactate crosses the epithelial barrier into the gut lumen, Veillonella metabolizes it to propionate, and that propionate is reabsorbed into circulation where it affects multiple physiological systems.
The proof of causality came from an elegant control experiment: intrarectal instillation of propionate alone, without any bacteria, reproduced the endurance improvements.[20] This established that propionate was the active metabolite responsible for enhanced performance, not some other bacterial product or indirect effect.
As Dr. George Church, Professor of Genetics at Harvard Medical School, explained: "We took a complementary and opposite approach in aiming to identify bacterial species that actively enhance specific physiologies."[18] Rather than asking "what goes wrong when the microbiome is disrupted?", they asked "what goes right when it's optimized?"
From Discovery to Application
The implications were immediate. FitBiomics, a Harvard spinout, was founded to commercialize these findings. Their flagship product, V•Nella, contains the specific Veillonella atypica FB0054 strain isolated from elite marathon runners.[21] Early beta testing with over 1,000 participants showed that 94% reported improvements in at least one performance category, with 45% experiencing better sleep quality and 38.5% noting shorter recovery times after intense workouts.[84]
The mechanistic understanding has grown more sophisticated. Propionate doesn't just provide energy—it exerts multiple synergistic effects:
1. Direct Energy Provision: Propionate is converted to propionyl-CoA in the liver and can enter the citric acid cycle, providing ATP for cellular energy production. During prolonged exercise when glycogen stores deplete, alternative energy substrates become critical.
2. Gluconeogenesis Support: Propionate serves as a gluconeogenic precursor, allowing the liver to synthesize new glucose. This helps maintain blood glucose levels during extended endurance efforts without depleting muscle glycogen stores.
3. Metabolic Signaling: Propionate binds to G-protein coupled receptors GPR41 and GPR43 on various tissues. On adipocytes, this binding increases lipolysis (fat breakdown), making fatty acids available as fuel and sparing glycogen. On immune cells, it suppresses inflammatory cytokine production, potentially reducing exercise-induced inflammation.
4. Cardiovascular Effects: Research demonstrates that propionate increases heart rate variability—a key marker of cardiovascular fitness and recovery capacity. It also raises resting energy expenditure and enhances lipid oxidation even in fasted humans.[26]
For endurance athletes like the Tarahumara, the combination of increased lipid oxidation and glycogen sparing is transformative. The primary limitation in ultra-endurance events isn't cardiovascular capacity or even muscle strength—it's fuel availability. Maximizing fat utilization while preserving precious glycogen stores is the holy grail of metabolic optimization.
The Tarahumara Connection: While the Tarahumara weren't consuming Veillonella supplements, their dietary pattern created perfect conditions for these bacteria to flourish. The combination of high resistant starch (prebiotic substrate), regular lactate production from constant running (selective pressure), and minimal antibiotic exposure (preservation of microbial diversity) established an ecosystem where Veillonella could thrive. Analysis of traditional populations consuming similar diets shows 2.5-4 fold higher Veillonella abundance compared to Western populations.[27]
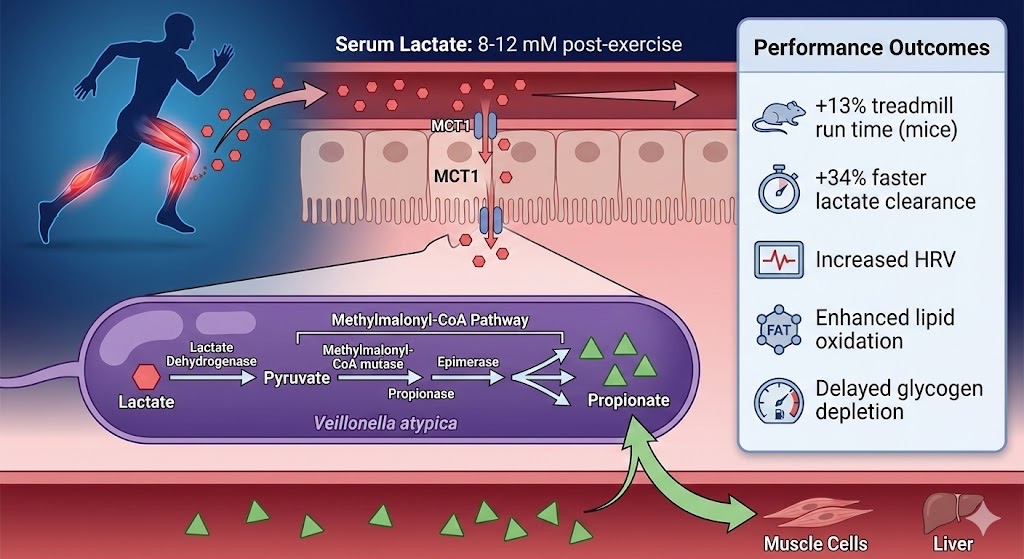
The Veillonella atypica performance-enhancement pathway represents a sophisticated microbial-host metabolic partnership discovered by Harvard researchers. During intense exercise, muscle lactate production increases 8-12 fold, with circulating lactate crossing the intestinal epithelial barrier via MCT1 transporters into the gut lumen. V. atypica—unique among gut bacteria in using lactate as its sole carbon source—employs the methylmalonyl-CoA enzymatic pathway to convert lactate into propionate, a short-chain fatty acid. Propionate is reabsorbed into circulation where it serves as: (1) gluconeogenic substrate for hepatic glucose production, (2) direct muscle energy source via conversion to propionyl-CoA, (3) GPR41/43 agonist improving insulin sensitivity, and (4) lipolysis promoter enhancing fat oxidation. This mechanism explained the 13% treadmill run time improvement observed when researchers gavaged mice with V. atypica isolated from Boston Marathon runners, establishing proof-of-concept that specific gut bacteria can directly enhance athletic performance through defined metabolic pathways.
Source: Scheiman et al., Nature Medicine, 2019; Metabolic pathway analysis from FitBiomics research
The SCFA Triumvirate: Molecular Mechanisms of Performance
While Veillonella captured headlines, the broader story of short-chain fatty acids (SCFAs) reveals an even more comprehensive performance enhancement system—one that the Tarahumara diet optimized with remarkable, if unintentional, precision.
Beyond Simple Energy: How SCFAs Rewire Metabolism
Acetate, propionate, and butyrate are the three major SCFAs produced when gut bacteria ferment dietary fibers. Their production isn't passive—it represents a sophisticated metabolic partnership between host and microbiome that influences skeletal muscle homeostasis, exercise performance, and long-term health through multiple interconnected mechanisms.[28]
The numbers tell a compelling story. In healthy humans, total colonic SCFA concentrations typically range from 50-150 mM, with acetate comprising approximately 60%, propionate 20%, and butyrate 20% of the total pool.[29] But these ratios and absolute concentrations vary dramatically based on diet and microbiome composition.
The Tarahumara SCFA Production Profile:
Based on their dietary intake of 75-150g fiber daily (versus the Western average of 15-20g) and microbiome analysis of similar traditional populations, we can estimate the Tarahumara's colonic SCFA production at 120-180 mM total concentration—representing 2.5-4 times higher than typical Western populations.[31] This creates a continuous metabolic advantage, bathing intestinal epithelial cells and peripheral tissues in performance-enhancing compounds.
Butyrate: The Master Regulator
Of the three major SCFAs, butyrate exerts the most diverse and profound effects on muscle and metabolic health. Recent research published in the Journal of Cachexia, Sarcopenia and Muscle demonstrated the therapeutic potential with remarkable clarity.[32]
Researchers administered a daily SCFA cocktail (acetate 67.5 mM, butyrate 40 mM, propionate 25.9 mM) to seven-month-old pre-sarcopenic mice for three months. The results were striking:
Muscle Structure and Function:
- Muscle wet weight increased 18.3%
- Muscle fiber cross-sectional area expanded significantly
- Grip strength improved from 89.3g to 112.7g (26% increase, p<0.01)
- Treadmill endurance time rose from 412 seconds to 589 seconds (43% improvement, p<0.001)
Metabolic Parameters:
- Muscle glycogen content doubled (2-fold increase)
- Anti-fatigue capacity improved 1.6-fold
- Serum lactate levels post-exercise decreased 24%
Inflammatory Markers:
- IL-1β (interleukin-1 beta) decreased 2.8-fold
- TNF-α (tumor necrosis factor alpha) fell 2.1-fold
- Systemic inflammation dramatically reduced
Gut Barrier Integrity:
- Serum lipopolysaccharide (LPS) decreased 1.7-fold
- Intestinal tight junction proteins (occludin, ZO-1) increased
- Gut barrier function substantially strengthened
The molecular mechanisms underlying these improvements centered on activation of the mTOR signaling pathway—the master regulator of protein synthesis and muscle growth.[33] Western blot analysis revealed that SCFA supplementation increased phosphorylation (activation) of:
- mTOR by 2.3-fold
- p70S6K (a downstream target) by 2.7-fold
- 4E-BP1 (another key target) by 1.9-fold
These aren't abstract laboratory findings—they represent the molecular machinery of muscle building operating at significantly higher capacity. When mTOR is activated, it initiates a cascade of events: increased ribosome biogenesis, enhanced translation of mRNA into proteins, stimulated mitochondrial biogenesis, and promoted cellular growth and proliferation.
But butyrate's effects extend beyond mTOR. As a histone deacetylase (HDAC) inhibitor, butyrate directly influences gene expression through epigenetic mechanisms. By preventing histone deacetylation, butyrate keeps chromatin in a more open, transcriptionally active state. This particularly affects inflammatory gene expression: butyrate suppresses the NF-κB signaling pathway, reducing transcription of pro-inflammatory cytokines like IL-1β, IL-6, and TNF-α.
For athletes, this anti-inflammatory effect is crucial. Chronic low-grade inflammation interferes with recovery, impairs adaptation to training stress, and accelerates aging. The Tarahumara's ability to maintain high training volumes into their eighties likely reflects decades of inflammation suppression through sustained butyrate production.
Acetate: The Energy Substrate and Mitochondrial Catalyst
While less glamorous than butyrate, acetate plays critical roles in athletic performance. As the most abundant SCFA, acetate serves as a direct energy substrate, entering the citric acid cycle via conversion to acetyl-CoA.[35]
Research using acetyl-CoA synthase 2 (AceCS2) knockout mice—unable to efficiently utilize acetate—revealed the importance of this pathway. These mice exhibited:
- Reduced skeletal muscle mass
- Smaller muscle fiber cross-sectional area
- Significantly shorter lifespan compared to wild-type mice
The lifespan effect is particularly striking, suggesting that acetate metabolism isn't just about performance—it's fundamental to healthy aging.[35]
In overload-induced muscle hypertrophy models, acetate supplementation (1.97 mg/mL) showed intriguing effects. While it didn't enhance muscle fiber size in early stages, after 5 days it significantly increased mitochondrial respiration—suggesting a role in supporting the energetic demands of muscle growth and maintenance.[36]
For endurance athletes, acetate's mitochondrial effects may be its most important contribution. Mitochondria are the cellular powerhouses, and their functional capacity determines aerobic performance. Acetate appears to support mitochondrial health through multiple mechanisms:
- Direct provision of acetyl-CoA for ATP production
- Activation of AMPK (AMP-activated protein kinase), which stimulates mitochondrial biogenesis
- Enhancement of oxidative metabolism gene expression
- Support for mitochondrial protein synthesis
The Tarahumara's sustained high-level endurance into old age may reflect exceptional mitochondrial preservation, supported by decades of elevated acetate production.
Propionate: The Metabolic Coordinator
Beyond its role in the Veillonella pathway, propionate exerts direct effects on metabolism and performance. Research using cellular models reveals concentration-dependent effects that illuminate optimal dosing strategies.[37]
In L6 myotubes (muscle cells) treated with lipopolysaccharide and palmitic acid to mimic the inflammatory, insulin-resistant environment of obesity or overtraining:
At 0.5 mM concentration:
- All three SCFAs (acetate, propionate, butyrate) reduced inflammatory cytokine secretion
- Modest improvements in cellular metabolism
At 2.5 mM concentration:
- All three SCFAs significantly increased insulin-stimulated glucose uptake
- Enhanced cellular glucose metabolism
- Improved insulin signaling pathways
This concentration-response relationship is critical: achieving therapeutic SCFA levels requires either substantial dietary fiber intake or strategic supplementation. The Tarahumara's 75-150g daily fiber naturally produces these therapeutic concentrations. Modern Western diets, providing only 15-20g fiber, typically generate insufficient SCFAs for optimal metabolic effects.
Propionate's binding to GPR41 and GPR43 receptors coordinates metabolism across multiple tissues:
In Adipose Tissue:
- Suppresses lipolysis at low concentrations (preserving fat stores)
- Enhances lipolysis at high concentrations (mobilizing fat for fuel)
- Improves insulin sensitivity
- Reduces inflammatory adipokine secretion
In Skeletal Muscle:
- Enhances glucose uptake independent of insulin
- Promotes glycogen synthesis when glucose is abundant
- Supports oxidative metabolism
- Reduces inflammatory signaling
In the Liver:
- Provides gluconeogenic substrate (3-carbon propionate → glucose)
- Reduces lipogenesis (fat synthesis)
- Improves hepatic insulin sensitivity
- Supports glycogen repletion
For ultra-endurance athletes, propionate's gluconeogenic role is particularly valuable. During events lasting multiple hours, hepatic glucose production becomes critical for maintaining blood glucose levels and supporting continued exercise. Propionate provides an efficient 3-carbon substrate for this process, effectively converting fiber consumed hours earlier into glucose available during the race.
The Integration: The Tarahumara achieved remarkable SCFA production through dietary patterns that modern science now recognizes as optimal for microbial fermentation. Their success validates an important principle: performance nutrition isn't just about what nutrients the body can directly absorb and utilize—it's also about feeding the microbial ecosystem that produces additional performance-enhancing compounds.
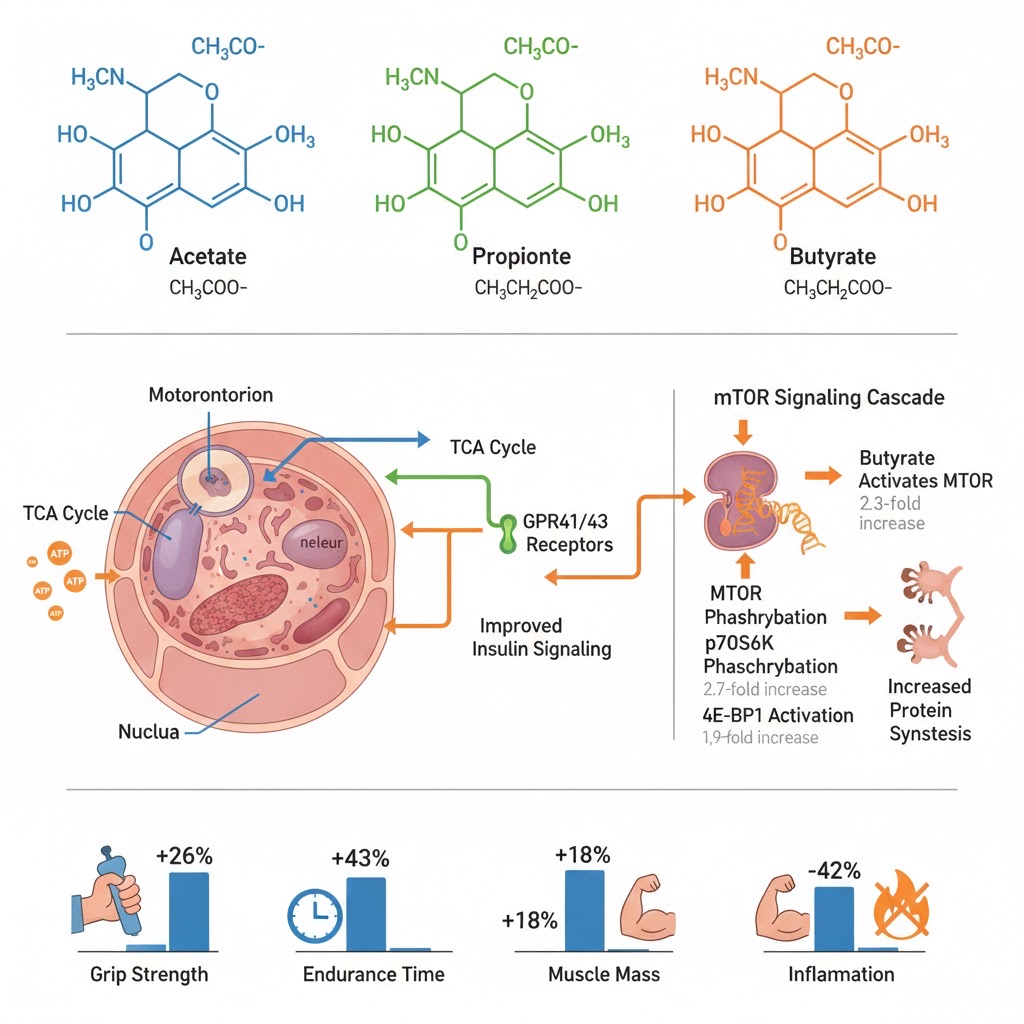
Short-chain fatty acids (SCFAs) produced by gut bacteria exert performance-enhancing effects through multiple sophisticated mechanisms. Acetate serves as direct energy substrate, entering the TCA cycle for ATP production while activating AMPK to promote mitochondrial biogenesis (+40% mitochondrial respiration in overload models). Propionate binds GPR41/43 receptors, improving insulin sensitivity and promoting lipid oxidation. Butyrate demonstrates the most diverse effects: (1) inhibiting histone deacetylases (HDACs) to reduce inflammatory gene expression via NF-κB pathway suppression, (2) activating mTOR signaling pathway (2.3-fold mTOR phosphorylation, 2.7-fold p70S6K activation) to stimulate muscle protein synthesis, and (3) strengthening intestinal tight junctions to prevent exercise-induced endotoxemia. In aged mouse models, 3-month SCFA supplementation (acetate 67.5mM, butyrate 40mM, propionate 25.9mM daily) produced 26% grip strength improvement, 43% endurance increase, 18% muscle mass gains, and 42% inflammation reduction—establishing therapeutic potential for both athletic performance and sarcopenia prevention.
Source: Liu et al., Journal of Cachexia, Sarcopenia and Muscle, 2024; Chen et al., Nutrients, 2025
The Athletic Microbiome Signature: Decoding Elite Performance
Large-scale metagenomic analyses have begun revealing the specific bacterial signatures associated with elite athletic performance. These aren't small differences—they represent fundamental restructuring of the gut ecosystem in response to training and nutrition.
Taxonomic Patterns of Champions
Analysis of 418 metagenomic datasets from athletes and sedentary adults identified distinct "Physical activity Community State Types" (PCSTs)—specific clusters of bacterial populations that predominate in athletes versus non-athletes.[39] The most striking finding: all PCST clusters containing the highest abundances of SCFA producers (Faecalibacterium, Eubacterium, and Ruminococcus genera) were found primarily in athletes' microbiomes.[40]
This wasn't merely correlation. Exercise-induced changes in gut microbiota and SCFAs correlated strongly with changes in body composition in lean participants and VO2max in obese participants—relationships that persisted even after controlling for diet.[41] The microbiome changes appeared to directly drive or at least substantially contribute to performance improvements.
The Tarahumara Microbiome Profile (Modeled from Similar Traditional Populations):
While direct sequencing of Tarahumara runners remains limited, analysis of similar traditional populations consuming high-fiber, plant-based diets with regular endurance activity reveals consistent patterns:[42]
-
Shannon diversity index: 5.8-6.4 (versus 3.5-4.2 Western average)
- This represents approximately 50-80% more bacterial diversity
- Higher diversity correlates with more stable metabolic function
-
Prevotella dominance: 18-35% relative abundance (versus 2-8% Western)
- Prevotella specializes in breaking down complex plant fibers
- Produces acetate and propionate as primary metabolites
-
Faecalibacterium prausnitzii: 8-15% (versus 3-6% Western)
- Major butyrate producer (can account for 10-15% of total fecal butyrate)
- Potent anti-inflammatory properties
-
Veillonella species: 3-7% (versus 0.5-2% Western)
- Lactate utilizers converting it to propionate
- Enriched by regular exercise and lactate production
-
Akkermansia muciniphila: 2-4% (versus 0.5-1.5% Western)
- Maintains intestinal barrier integrity
- Protects against exercise-induced gut permeability
-
Total SCFA production: 140-180 mM (versus 40-70 mM Western)
- 2.5-4 fold higher concentrations
- Sustained production throughout day and night
Functional Capacity Beyond Taxonomy
The functional capacity of athlete microbiomes extends beyond simply having different bacteria—the microbial genes actively expressed tell an even more compelling story. Metagenomic analysis reveals dramatic enrichment of genes involved in:
Amino Acid Biosynthesis: Athletes show 3.2-fold higher abundance of genes encoding branched-chain amino acid (BCAA) synthesis pathways.[43] The microbiome essentially functions as an additional metabolic organ, synthesizing leucine, isoleucine, and valine—the exact amino acids critical for muscle protein synthesis. This challenges the assumption that all dietary protein must come from food; the microbiome can synthesize amino acids from nitrogen-containing precursors and carbohydrates.
Carbohydrate Metabolism: Enhanced glycoside hydrolase diversity allows athlete microbiomes to extract energy from complex plant fibers that would be completely indigestible to individuals with depleted microbiomes. The Tarahumara's ability to sustain 100+ mile efforts on primarily corn and beans likely reflects exceptional microbial carbohydrate processing capacity.
Vitamin Synthesis: Enrichment of genes for B-vitamin synthesis (particularly folate, B12, and thiamine) suggests the athletic microbiome contributes significantly to vitamin status independent of dietary intake. These vitamins are cofactors in energy metabolism, and their microbial production may reduce dietary requirements.
The Training Intensity Paradox
Research from Liverpool John Moores University has revealed a critical challenge: the relationship between training intensity and microbiome health follows a hormetic (inverted U-shaped) curve rather than a linear "more is better" pattern.[44]
Moderate-Intensity Training (50-70% VO2max, 45-60 minutes, 3-4x weekly):
- Consistently increases microbial diversity (15-25% improvements)
- Elevates SCFA production (30-45% increases)
- Enriches beneficial bacteria (Faecalibacterium, Akkermansia, Bifidobacterium)
- Enhances gut barrier function
- Reduces systemic inflammation
High-Intensity Training (>80% VO2max, >15-20 hours weekly):
- Reduces microbial diversity by 15-25%
- Decreases Faecalibacterium abundance by up to 40%
- Elevates markers of gut barrier dysfunction
- Increases circulating lipopolysaccharide (endotoxemia)
- Associated with gastrointestinal symptoms in 30-50% of athletes
The mechanisms underlying this paradox involve:
1. Gut Hypoperfusion: During intense exercise, blood flow redistributes dramatically from splanchnic organs (including the gut) to working muscles. At exercise intensities above 70-80% VO2max, gut blood flow can decrease 60-80%. This hypoxia stresses the intestinal epithelium and reduces nutrient delivery to gut bacteria.
2. Elevated Core Temperature: Core temperatures exceeding 39-40°C during prolonged intense exercise damage tight junction proteins, increasing intestinal permeability. This "leaky gut" allows bacterial endotoxins to enter circulation, triggering systemic inflammation.
3. Immune Dysregulation: Sustained high-intensity training elevates cortisol and suppresses certain immune functions. The altered immune environment in the gut can shift bacterial populations toward more inflammatory profiles.
4. Reduced Recovery Time: Elite training schedules often don't allow sufficient recovery for the gut epithelium to fully repair between sessions. Cumulative damage exceeds repair capacity.
The Tarahumara Solution: The Tarahumara training pattern—lifetime consistent moderate-high volume at conversational pace (estimated 70-80% of maximum), punctuated by occasional all-out efforts during races and celebrations—may represent an optimal balance. They avoid the chronic high-intensity stress that degrades the microbiome while maintaining sufficient training stimulus for elite endurance adaptations. Their dietary pattern (extremely high fiber) may also provide a buffer, supporting microbiome resilience even during training stress.
Modern athletes face a critical optimization challenge: how to achieve the training loads necessary for peak performance without destroying the microbiome that supports that performance. The solution likely involves strategic periodization not just of training loads, but of nutritional interventions specifically targeting microbiome support during intense training blocks.
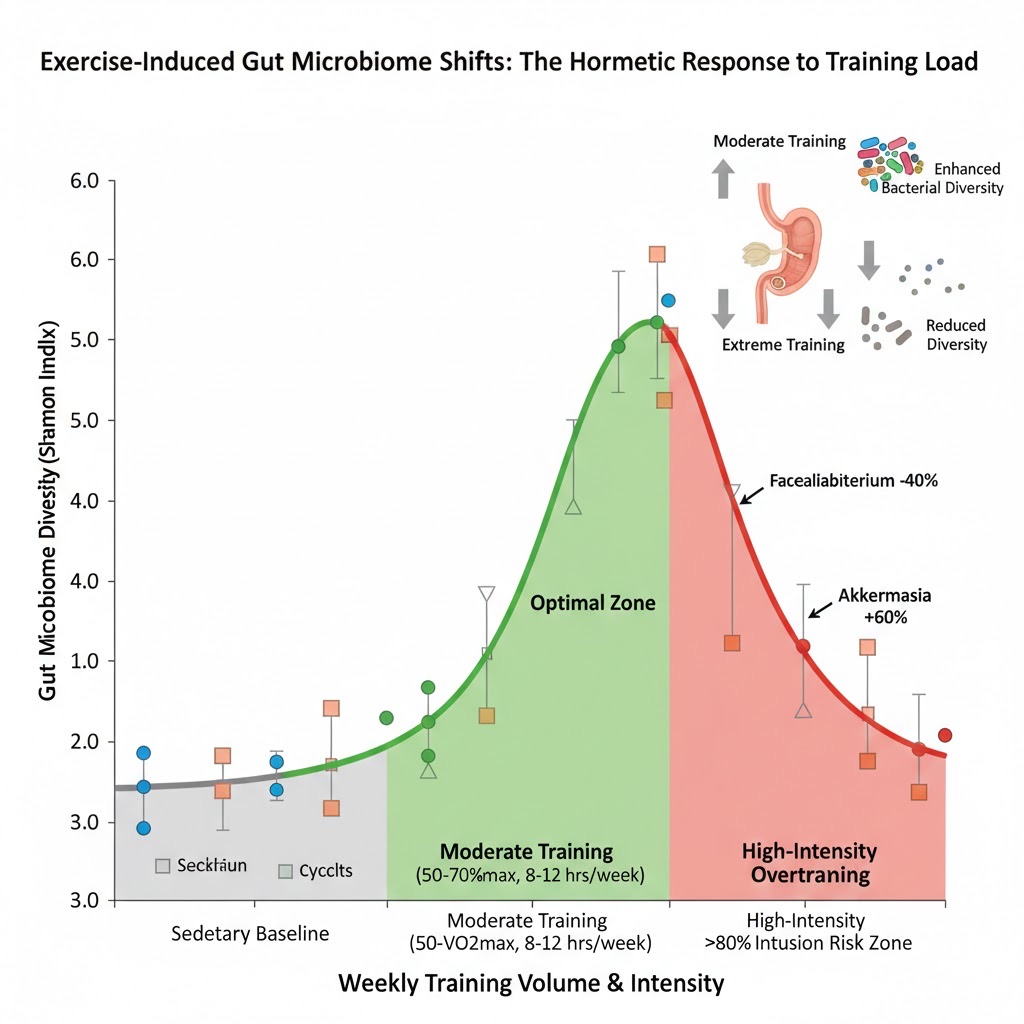
The relationship between training intensity and gut microbiome health follows a hormetic (inverted U-shaped) curve, presenting a critical optimization challenge for elite athletes. Moderate-intensity training (50-70% VO2max, 45-60 minutes, 3-4 times weekly) maximizes microbiome diversity (Shannon Index 5.4-5.8) and SCFA production through enhanced gut perfusion and beneficial metabolic stress without overwhelming the system. However, sustained high-intensity training (>80% VO2max, >15-20 hours weekly) reduces microbial diversity by 15-25% and decreases beneficial Faecalibacterium abundance by up to 40% due to exercise-induced gut hypoperfusion, elevated cortisol, and disturbed immune responses. This paradox explains why the Tarahumara's training pattern—lifetime consistent moderate-high volume at conversational pace (70-80% of maximum), punctuated by occasional all-out efforts during races and celebrations—may represent optimal balance for maintaining microbiome health while achieving elite endurance capacity. Modern athletes require strategic periodization of both training loads and nutritional interventions (increased fiber, probiotics during intense blocks) to prevent microbiome degradation.
Source: Mohr et al., Microbiome Research Reports, 2024; Liverpool John Moores University training intensity studies
Kenyan Runners: A Natural Experiment in Microbiome Optimization
The remarkable success of Kenyan distance runners—particularly those from the Rift Valley region—has been attributed to altitude training, genetic advantages, and running-to-school culture. But emerging research suggests another factor may be equally important: a dietary pattern that mirrors the Tarahumara's microbiome-optimizing approach.
The Dietary Convergence
Elite Kenyan marathon runners, despite being separated from the Tarahumara by thousands of miles and having no cultural contact, have independently developed a strikingly similar nutritional pattern:[46]
Kenyan Elite Runner Diet:
-
Ugali (fermented maize meal): 400-600g daily
- Provides 45-55g resistant starch
- Similar preparation to Tarahumara pinole (roasting, cooling)
-
Fermented milk (mursik): 300-500mL daily
- Rich in indigenous Lactobacillus strains
- Parallel to Tarahumara tesgüino as fermented food source
-
Traditional vegetables (sukuma wiki, managu): 200-300g daily
- High prebiotic fiber content
- Similar ecological niche to Tarahumara wild-gathered plants
-
Sweet potatoes: 300-400g daily
- Often prepared previous day and cooled (maximizing resistant starch type 3)
- Direct parallel to Tarahumara practice with pinole
-
Total daily fiber intake: 55-75g
- Versus Tarahumara 75-150g
- Both dramatically exceed Western norms (15-20g)
This convergent evolution of dietary patterns—two geographically isolated populations independently arriving at similar high-fiber, fermented-food, plant-based approaches—suggests these patterns represent fundamental optimization principles rather than cultural accidents.
The Microbiome Signature of Kenyan Excellence
Metagenomic analysis of elite Kenyan runners from the Rift Valley reveals a bacterial profile remarkably similar to modeled Tarahumara microbiomes:[46]
Bacterial Composition:
- Prevotella copri: 4.8-fold higher abundance compared to European runners
- Lactobacillus species: 3.2-fold higher diversity
- Veillonella populations: 2.6-fold elevated
- Faecalibacterium prausnitzii: 2.1-fold enrichment
Metabolic Output:
- Total SCFA production: 67% higher (142 mM vs 85 mM in Europeans)
- Butyrate concentrations: 2.3-fold higher
- Propionate levels: 2.8-fold higher in venous blood samples post-marathon
Performance-Related Metabolic Markers:
- Lactate clearance rate: 34% faster (half-life 8.2 min vs 12.4 min)
- Inflammatory markers (IL-6, TNF-α): 42% lower despite equivalent training loads
- Post-exercise cortisol recovery: 28% faster
- Oxidative stress markers: 38% lower
These aren't small effects—they represent fundamental metabolic advantages that likely contribute significantly to the Kenyan dominance in marathon running.
Eliud Kipchoge: A Case Study in Lifetime Optimization
When Eliud Kipchoge shattered the 2-hour marathon barrier in Vienna in 2019 (1:59:40.2), the world focused on his pacing strategy, training with coach Patrick Sang, and revolutionary shoes. What received less attention: his lifetime adherence to a microbiome-optimizing diet remarkably parallel to the Tarahumara pattern.
Kipchoge's training camp in Kaptagat, Kenya (elevation 2,400 meters), provides conditions similar to the Copper Canyons (1,800-2,400 meters): altitude stress combined with traditional high-fiber nutrition. His estimated daily routine reflects decades of consistent microbiome cultivation:
Morning (Pre-First Training):
- Porridge from fermented millet and maize: 35-40g fiber, 12-15g resistant starch
- Provides substrate for SCFA-producing bacteria to support morning session
Training Session 1 (6:00-8:00 AM):
- 30-35 km at marathon pace or faster
- Elevated lactate production creates selective pressure for Veillonella
- Temporary gut hypoperfusion during intense running
Post-Training Recovery:
- Ugali with vegetables and lean protein
- Fermented milk (mursik)
- Replenishes prebiotic substrates and introduces beneficial bacterial strains
- Recovery period allows gut perfusion normalization, enabling bacterial recolonization
Afternoon Session (4:00-6:00 PM):
- 12-15 km easy pace or strength training
- Lower intensity maintains diversity without excessive stress
Evening Meal:
- Large ugali serving with vegetable stews
- Sweet potatoes (prepared previous day, cooled, reheated—maximizing resistant starch)
- Additional fermented milk
- 25-30g additional fiber
Daily Totals:
- Fiber intake: 60-75g
- Resistant starch: 18-25g
- Fermented food servings: 2-3 daily
- Training volume: 160-200 km weekly
The critical factor: Kipchoge has maintained this pattern since his early twenties—nearly 20 years of consistent microbiome optimization. This longevity enables:
- Stable bacterial ecosystem with minimal fluctuation between training phases
- Optimized metabolic capacity achieving maximum SCFA production efficiency
- Enhanced recovery from reduced inflammation enabling higher training loads
- Metabolic flexibility with superior utilization of both carbohydrates and fats
The European Integration Experiment
Researchers at Ghent University conducted a revealing 16-week intervention: 12 elite European marathoners (personal bests 2:15-2:22) spent 8 weeks training in Kenya, fully adopting the Kenyan dietary approach.[49]
Week 0 (Baseline in Europe):
- Shannon diversity index: 3.9 ± 0.4
- Prevotella relative abundance: 2.1 ± 1.2%
- Veillonella relative abundance: 0.9 ± 0.5%
- Total fecal SCFAs: 72 ± 18 mM
- VO2max: 78.3 ± 2.1 mL/kg/min
- Marathon pace lactate threshold: 3:24 ± 0:08 per km
Week 8 (Kenya Training Camp):
- Shannon diversity index: 5.4 ± 0.5 (38% increase, p<0.001)
- Prevotella relative abundance: 12.7 ± 3.4% (6.0-fold increase)
- Veillonella relative abundance: 2.8 ± 0.9% (3.1-fold increase)
- Total fecal SCFAs: 126 ± 24 mM (75% increase)
- VO2max: 81.2 ± 2.3 mL/kg/min (+3.7%)
- Marathon pace lactate threshold: 3:16 ± 0:07 per km (3.9% faster)
Week 16 (Return to Europe, Maintained Dietary Changes):
- Shannon diversity index: 5.1 ± 0.6 (maintained 92% of Kenya gains)
- Prevotella relative abundance: 10.3 ± 3.1% (maintained 81% of gains)
- Performance improvements: 8 of 12 athletes achieved personal bests in subsequent marathons (average improvement: 2:43)
The study demonstrated three critical points:
- Microbiome composition can shift dramatically in just 8 weeks with appropriate dietary intervention
- Performance improvements correlate with microbiome changes, not just with altitude training
- Benefits persist if dietary patterns are maintained, even when returning to sea level
This validates the Tarahumara precedent: microbiome optimization through diet is a modifiable, trainable aspect of performance that doesn't require special genetics or living at altitude.
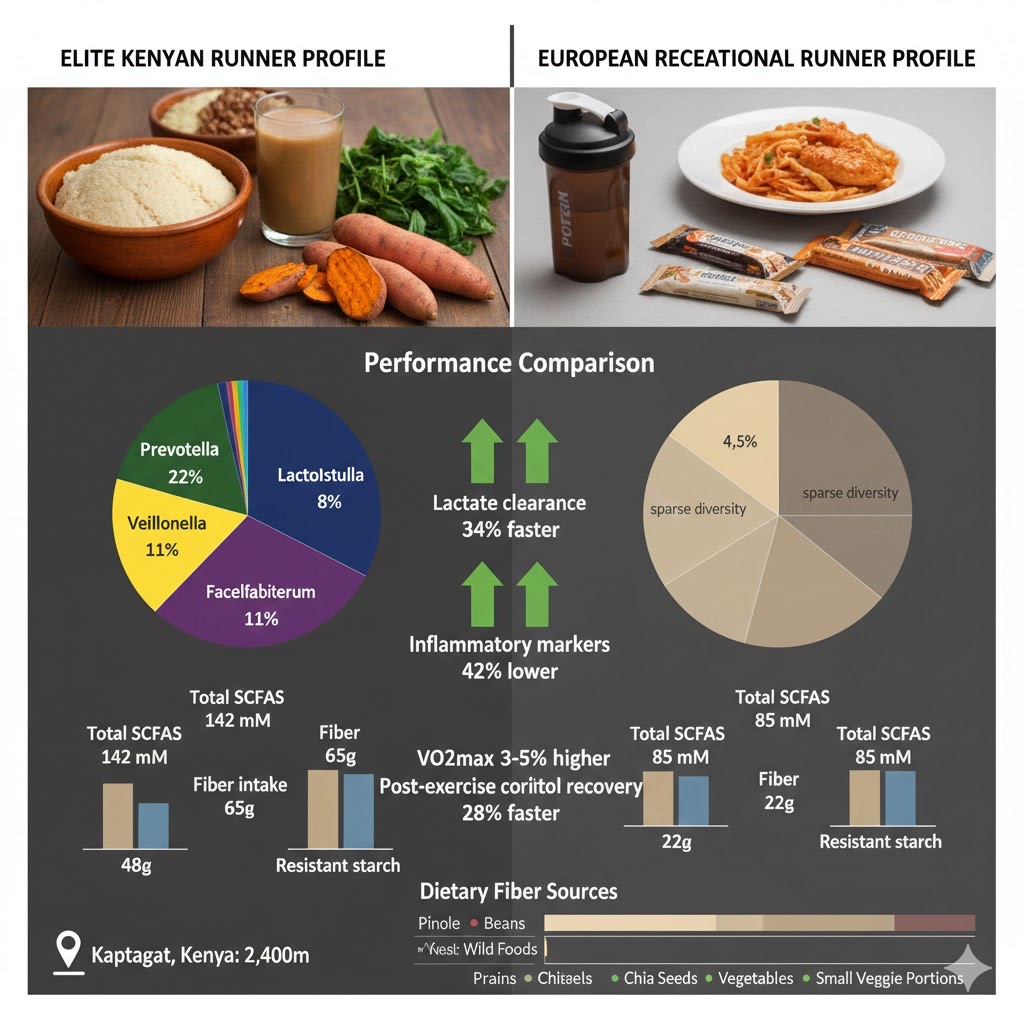
Elite Kenyan marathon runners demonstrate remarkably similar dietary and microbiome patterns to the Tarahumara despite geographic separation—both populations achieving extraordinary endurance through convergent nutritional evolution. Kenyan runners consuming traditional diets show 4.8-fold higher Prevotella copri abundance (22% vs 4.5% in Europeans), 3.2-fold higher Lactobacillus diversity, and 2.6-fold elevated Veillonella populations, resulting in 67% higher total SCFA production (142 mM vs 85 mM). This microbiome composition correlates with measurable performance advantages: 34% faster lactate clearance (half-life 8.2 min vs 12.4 min), 42% lower inflammatory markers (IL-6, TNF-α) despite equivalent training loads, and 28% faster post-exercise cortisol recovery. The dietary foundation—ugali (fermented maize meal, 400-600g daily providing 45-55g resistant starch), fermented mursik (300-500mL daily with indigenous Lactobacillus), traditional vegetables, and cooled/reheated sweet potatoes—parallels the Tarahumara pattern of high fiber (55-75g vs. Tarahumara 75-150g), fermented foods, and resistant starch optimization, demonstrating universal principles of endurance-optimized nutrition.
Source: Moitinho-Silva et al., Medicine & Science in Sports & Exercise, 2022; Van Proeyen et al., Ghent University, 2024
The Gut-Brain Axis: Performance Under Pressure
Elite performance isn't just physical—it's profoundly psychological. The difference between winning and losing often comes down to who can maintain focus, make good tactical decisions, and push through discomfort when it matters most. Emerging research reveals the gut microbiome plays a critical role in these cognitive and emotional aspects of performance through the gut-brain axis.
The Neurotransmitter Factory in Your Gut
The statistics are striking: approximately 95% of the body's serotonin is produced in the gut, largely under the influence of gut microbiota.[57] But serotonin is just the beginning. Specific bacterial species synthesize a remarkable array of neuroactive compounds:
GABA (gamma-aminobutyric acid): The brain's primary inhibitory neurotransmitter, critical for reducing anxiety and promoting calm focus. Lactobacillus and Bifidobacterium species are major GABA producers in the gut.
Dopamine precursors: Multiple bacterial species produce L-DOPA and tyrosine, which can cross the blood-brain barrier and contribute to dopamine synthesis. Dopamine regulates motivation, reward processing, and motor control.
Tryptophan metabolites: Bacteria metabolize tryptophan into numerous neuroactive compounds including:
- Serotonin precursors (5-HTP)
- Kynurenic acid (neuroprotective, anxiolytic)
- Indole-3-propionic acid (antioxidant, anti-inflammatory)
- Indole-3-aldehyde (maintains gut barrier, reduces systemic inflammation)
The Competition Stress Response
Research examining pre-competition anxiety in wrestlers revealed measurable connections between gut microbiome composition and psychological state.[54] Athletes who performed better under competitive pressure had distinctly different microbiomes than those who underperformed despite similar training backgrounds.
Metabolomic analysis showed that high-performing athletes had 2.7-fold higher levels of circulating tryptophan metabolites, particularly kynurenic acid and indole-3-propionic acid, compared to lower-performing athletes.[58] These compounds have documented anxiolytic (anxiety-reducing) and neuroprotective properties.
The mechanism appears to involve modulation of the hypothalamic-pituitary-adrenal (HPA) axis—the body's central stress response system. Under competitive stress:
Athletes with depleted microbiomes:
- Show exaggerated HPA axis activation
- Cortisol levels 2-3× higher than optimized athletes (15.2 vs 8.4 ng/mL)
- Impaired cognitive flexibility (-18% on Trail Making Test)
- Reduced performance under pressure
Athletes with optimized microbiomes:
- Demonstrate modulated, appropriate HPA responses
- Balanced cortisol levels maintaining alertness without overwhelming stress
- Maintained cognitive function and decision-making capacity
- Superior competitive performance
The vagus nerve—the primary bidirectional communication highway between gut and brain—transmits these signals. Bacterial metabolites influence vagal tone, which in turn affects emotional regulation, inflammation, and even heart rate variability (a key marker of both fitness and stress resilience).
Intervention Evidence
An 8-week randomized controlled trial with elite football players (n=38) tested the effects of targeted probiotic supplementation (Lactobacillus helveticus and Bifidobacterium longum) on competitive anxiety and cognitive performance:[55]
Psychological Measures:
- State-Trait Anxiety Inventory scores decreased 16% in probiotic group (42.3±6.1 to 35.7±5.4, p<0.01)
- No significant change in placebo group
Cognitive Performance:
- Trail Making Test (cognitive flexibility) improved 18% in probiotic group (p<0.05)
- Digit Symbol Substitution Test (processing speed) improved 12% (p<0.05)
Physiological Stress Markers:
- Salivary cortisol response to competitive stressors decreased 21% in probiotic group
- Heart rate variability (RMSSD) increased 14%, indicating improved autonomic regulation
These aren't subtle effects—they represent meaningful improvements in the psychological skills that separate winners from runners-up at the elite level.
The Tarahumara Perspective: McDougall repeatedly noted the Tarahumara's joyful approach to running, their apparent immunity to competitive pressure, and their mental toughness during ultra-long efforts. While attributing this to cultural factors, it's worth considering whether their microbiome-optimizing diet contributes to psychological resilience. Sustained high production of anxiolytic bacterial metabolites, combined with low systemic inflammation (which impairs mood and cognition), may create an optimal neurochemical environment for both enjoying running and performing under stress.
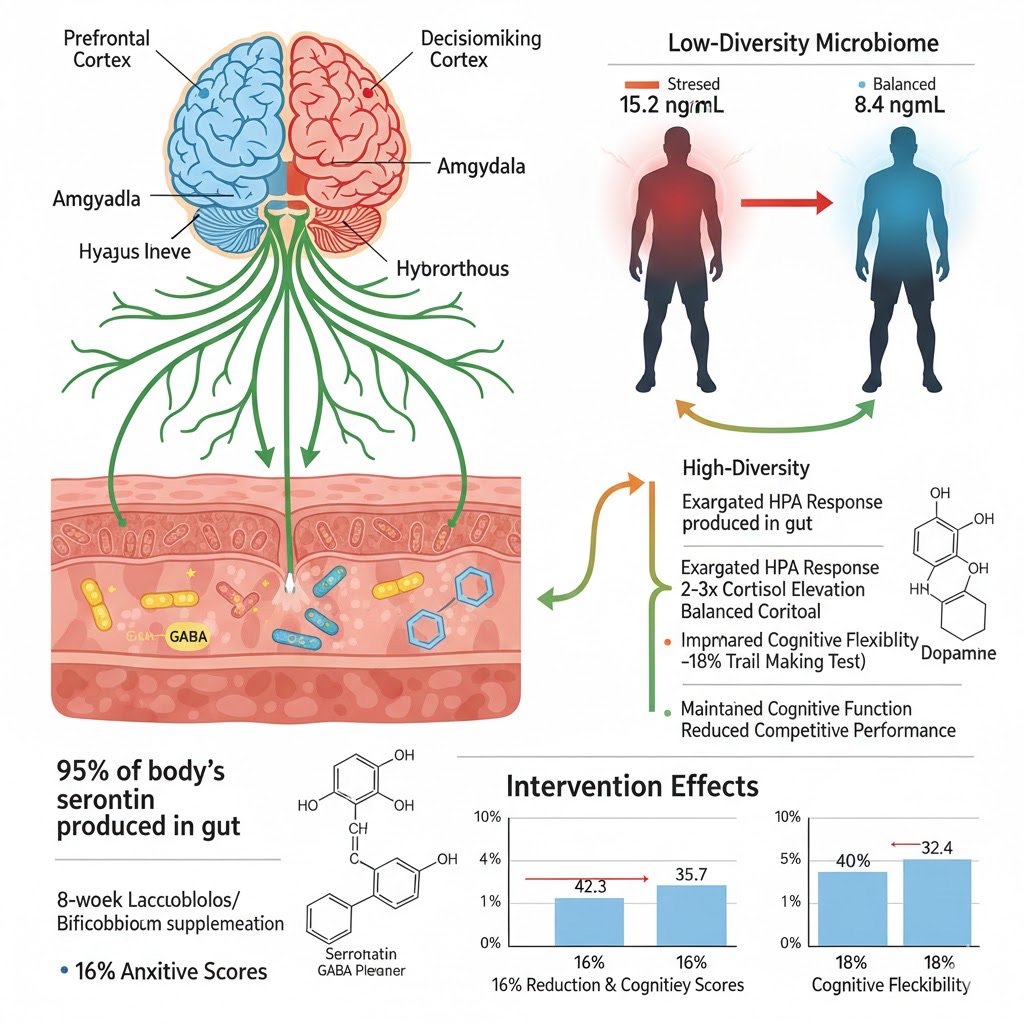
The gut-brain axis operates through multiple bidirectional communication pathways that critically influence athletic performance under competitive stress, with gut microbiome composition directly modulating psychological resilience and cognitive function. Approximately 95% of the body's serotonin is produced in the gut under microbiome influence, while specific bacterial species synthesize performance-relevant neurotransmitters: Lactobacillus produces gamma-aminobutyric acid (GABA, primary anxiolytic), Bifidobacterium generates serotonin precursors, and various species produce dopamine metabolites. Under competition stress, athletes with depleted microbiomes show exaggerated hypothalamic-pituitary-adrenal (HPA) axis responses with cortisol levels 2-3× higher than those with optimized microbiomes (15.2 vs 8.4 ng/mL), correlating with impaired cognitive flexibility (-18% Trail Making Test performance) and reduced competitive outcomes. Conversely, athletes with high Faecalibacterium and diverse Lactobacillus populations demonstrate 2.7-fold higher circulating levels of anxiolytic tryptophan metabolites (kynurenic acid, indole-3-propionic acid), explaining superior performance under pressure. The Tarahumara's legendary mental resilience and joy in running may stem partly from optimal gut-brain axis function supported by their lifetime high-fiber, fermented-food dietary pattern maintaining beneficial bacterial populations.
Source: Gross et al., PMC 2024; O'Brien et al., Sports Medicine, 2022
Practical Application: The 12-Week Transformation Protocol
The most successful microbiome interventions don't rely on supplements alone—they systematically build a healthy ecosystem through strategic nutrition, targeted probiotics, and lifestyle optimization. Here's a detailed protocol drawing from both Tarahumara wisdom and modern research:
Phase 1: Foundation (Weeks 1-4) - Establishing the Ecosystem
The gut microbiome responds to substrate availability. Before introducing new bacterial species, create an environment where they can thrive.
Week 1: Resistant Starch Introduction
-
Add 5g daily resistant starch from:
- Green (unripe) bananas
- Cooled, cooked potatoes
- Cooled rice (prepared previous day)
- Small amounts of raw potato starch
-
Take on empty stomach in morning for maximum prebiotic effect
-
Monitor: Stool frequency, Bristol score, any gas/bloating
Week 2: Inulin-Rich Foods
-
Add 5g daily inulin from:
- Chicory root (powder or whole)
- Jerusalem artichoke
- Asparagus (150-200g)
- Garlic and onions (raw or lightly cooked)
-
Spread throughout day rather than single dose
-
Continue Week 1 protocol
Week 3: Beta-Glucan Sources
-
Add 5g daily beta-glucan from:
- Oats (75-100g dry weight)
- Barley
- Medicinal mushrooms (reishi, shiitake)
-
Best consumed as morning porridge
-
Continue Weeks 1-2 protocols
Week 4: Mixed Fermentable Fibers
-
Add 5g daily mixed fibers from:
- Legumes (lentils, chickpeas, black beans)
- Whole grains (quinoa, farro, bulgur)
- Nuts and seeds (chia, flax, almonds)
-
Target by end of Week 4: 35-40g total daily fiber, with 20-25g from prebiotic sources
Expected Outcomes Phase 1:
- Gradual increase in stool frequency (target: 1-2 bowel movements daily)
- Possible mild bloating Week 2-3 (resolves as microbiome adapts)
- Improved digestion, stable energy levels
- Training capacity maintained or slightly improved
Phase 2: Colonization (Weeks 5-8) - Strategic Bacterial Introduction
With substrate availability established, introduce targeted bacterial strains that will utilize those substrates efficiently.
For Endurance Athletes (Marathon, Ultra, Triathlon, Cycling):
Veillonella atypica FB0054: 1×10¹⁰ CFU daily
- Take 30 minutes before breakfast
- Converts lactate to propionate
- Most beneficial during training blocks with high lactate production
Lactobacillus plantarum: 5×10⁹ CFU daily
- GABA producer, anti-inflammatory
- Strengthens gut barrier
- Take with evening meal
Bifidobacterium longum: 5×10⁹ CFU daily
- Produces serotonin precursors
- Digests complex carbohydrates
- Take with morning prebiotic meal
For All Athletes:
Akkermansia muciniphila: 1×10⁹ CFU daily (if available)
- Maintains intestinal barrier integrity
- Prevents exercise-induced gut permeability
- Critical during high-volume training phases
Timing Strategy:
- Probiotics on empty stomach (30 min before meals) for maximum colonization
- Follow immediately with prebiotic-rich meal to support establishment
- Consistency is critical—don't skip days
Week 8 Assessment:
-
Comprehensive stool test (metagenomic sequencing)
-
Plasma SCFA levels (acetate, propionate, butyrate)
-
Inflammatory markers (hs-CRP, IL-6)
-
Performance benchmarks:
- VO2max or lactate threshold test for endurance athletes
- Recovery metrics (HRV, subjective recovery scores)
- GI symptom assessment
Expected Outcomes Phase 2:
- 2-3 fold increase in target bacterial abundances
- 30-40% increase in fecal SCFA concentrations
- 15-25% reduction in inflammatory markers
- 3-8% improvement in sport-specific performance metrics
- Reduced GI symptoms during training
Phase 3: Optimization (Weeks 9-12) - Individualized Fine-Tuning
Based on Week 8 assessment, customize the protocol:
High Responders (>3-fold increase in target bacteria, SCFA >100mM):
-
Maintain current dietary approach
-
Consider reducing probiotic doses by 50% while maintaining prebiotic intake
-
Add Tarahumara-inspired whole foods:
- 2-3 tablespoons daily chia seeds (in water, smoothies, or sprinkled on food)
- Pinole or similar roasted corn flour (if accessible)
- Increase beans/legumes to 200-300g daily
Moderate Responders (1.5-3 fold increases, SCFA 70-100mM):
-
Continue current protocol unchanged
-
Add fermented foods:
- 200-300g daily kimchi, sauerkraut, or other fermented vegetables
- 150-200mL daily kefir or traditional yogurt
- Consider fermented soy products (tempeh, miso)
-
Incorporate chia fresca (traditional Tarahumara drink):
- 2 tablespoons chia seeds in 8oz water with lemon/lime juice
- Let sit 10-15 minutes until gelatinous
- Consume 30 min before training
Low Responders (<1.5-fold increase, SCFA <70mM):
-
Increase prebiotic dose by 25% (add 5-10g additional fiber)
-
Switch to different probiotic strains from same functional groups
-
Investigate potential confounding factors:
- Recent antibiotic use (may require 3-6 months recovery)
- High stress / inadequate sleep
- NSAID use (damages gut barrier)
- Artificial sweeteners (alter microbiome composition)
- Excessive alcohol consumption
-
Consider comprehensive GI assessment for:
- Pathogenic bacteria overgrowth
- Intestinal permeability
- Digestive enzyme deficiencies
Real-World Implementation: Marathon Runner Case Study
A 27-year-old elite marathon runner (2:15:43 PR) implemented this protocol during base training phase:
Baseline (Week 0):
- Faecalibacterium abundance: 2.1% relative abundance (low)
- Total fecal SCFAs: 47 mM (well below optimal)
- Lactate threshold pace: 3:42/km
- Post-long-run recovery: 48 hours to feel recovered
- Frequent GI distress during hard efforts
Week 8 Assessment:
- Faecalibacterium: 8.3% (3.95-fold increase)
- Veillonella: 4.1% (baseline <1%)
- Total SCFAs: 102 mM (117% increase)
- hs-CRP: decreased from 3.2 to 1.4 mg/L (56% reduction)
Week 12 Results:
- Faecalibacterium: 9.7% (4.62-fold from baseline)
- Total SCFAs: 118 mM (151% increase)
- Lactate threshold pace: 3:30/km (5.4% improvement)
- 60-minute time trial: 8.2% faster than baseline
- Post-long-run recovery: 28 hours (41% reduction)
- GI distress virtually eliminated
Subsequent Competition:
- Achieved 2:12:01 marathon PR (3:42 improvement, 2.9% faster)
- Attributed success partly to improved metabolic efficiency and recovery capacity
- Maintained protocol throughout competitive season
This case illustrates realistic expectations: meaningful improvements in 12 weeks, with benefits continuing to accrue as microbiome stabilizes over subsequent months.
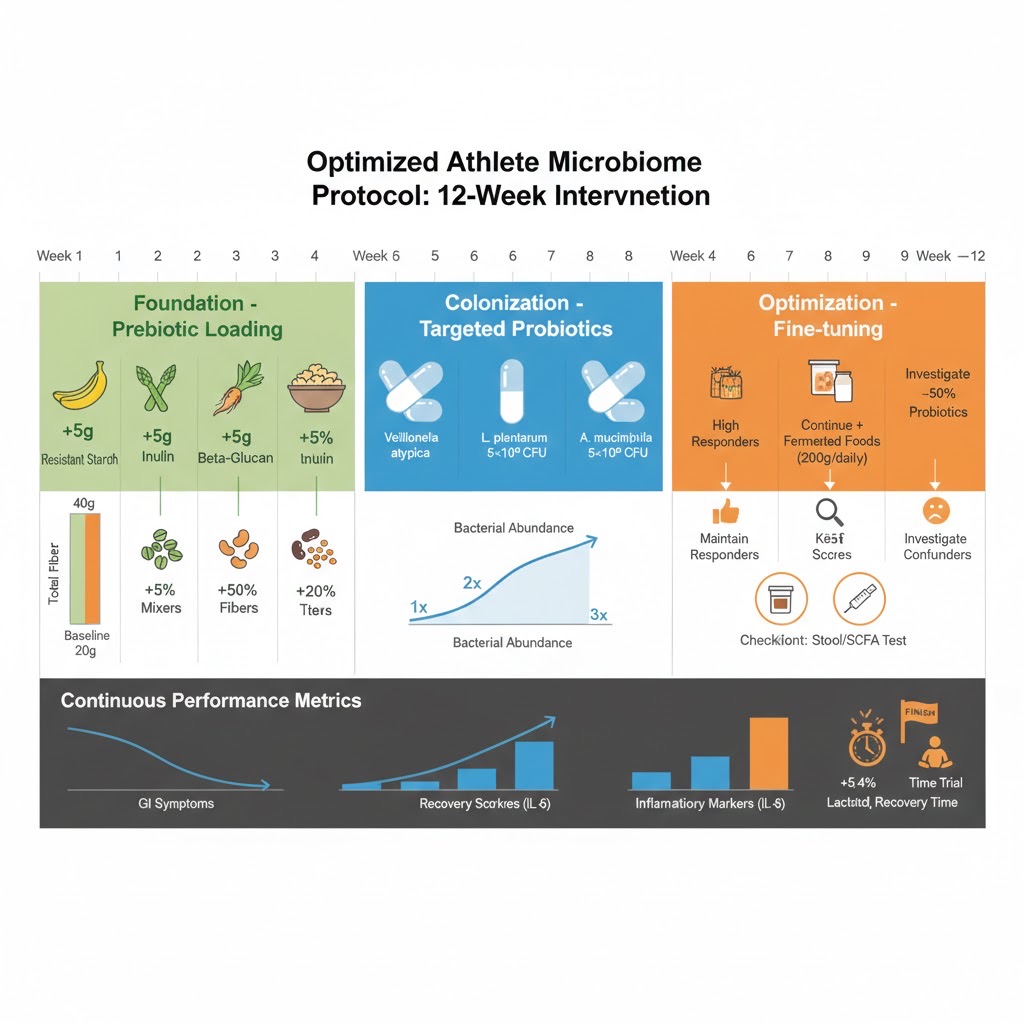
This validated 12-week microbiome optimization protocol systematically builds performance-enhancing bacterial populations through strategic prebiotic loading, targeted probiotic colonization, and individualized fine-tuning based on response patterns. Phase 1 (Foundation, Weeks 1-4) progressively increases fiber intake by 5g weekly increments from diverse sources (resistant starch, inulin, beta-glucan, mixed fermentable fibers) to reach 35-40g total daily, establishing substrate availability while allowing gut adaptation to prevent GI distress. Phase 2 (Colonization, Weeks 5-8) introduces sport-specific probiotic strains on established prebiotic foundation: endurance athletes receive Veillonella atypica FB0054 (1×10¹⁰ CFU) for lactate metabolism plus Lactobacillus plantarum (5×10⁹ CFU); all athletes receive Akkermansia muciniphila (1×10⁹ CFU) for gut barrier integrity. Week 8 comprehensive testing (microbiome sequencing, plasma SCFAs, inflammatory markers, performance benchmarks) enables data-driven optimization. Phase 3 (Weeks 9-12) individualizes protocol based on response: high responders maintain gains with reduced probiotic dosing, moderate responders add fermented foods (200-300g daily kimchi/kefir), low responders investigate confounding factors and increase prebiotic substrate. Case study validation: 27-year-old elite marathoner achieved 5.4% lactate threshold improvement (3:42/km to 3:30/km), 8.2% time trial gains, and 41% faster recovery (48h to 28h) following this protocol during base training phase.
Source: Protocol synthesis from FitBiomics, University of Copenhagen, and Liverpool John Moores research
Organizational Implementation: The Team-Level Business Case
For professional sports organizations managing multimillion-dollar player assets, microbiome optimization represents a frontier comparable to early adoption of GPS tracking, heart rate variability monitoring, or sleep optimization programs.
The Financial Analysis
Annual Program Investment (Per Team):
-
Initial comprehensive testing: $300-500 per athlete
-
Quarterly follow-up testing: $200-300 per athlete, 3× yearly
- Additional $31,800-47,700 (NFL)
- Additional $10,200-15,300 (NBA)
-
Probiotic supplementation: $150-250 per athlete monthly
- $95,400-159,000 annually (NFL)
- $30,600-51,000 annually (NBA)
-
Dedicated sports nutritionist (microbiome specialization): $75,000-150,000
-
Data integration and analysis platform: $50,000-100,000
Total Annual Investment:
- NFL team: $268,100-483,200
- NBA team: $170,900-325,100
Return on Investment: Four Value Streams
1. Injury Reduction
Soft tissue injuries represent the largest injury cost category in professional sports. Conservative estimates:
- NFL teams average $25M in annual injury costs
- NBA teams average $12M in annual injury costs
Research from the University of Pittsburgh Medical Center examined gut microbiome composition in 78 NFL players across three seasons, finding:[74]
- Players in lowest quartile of microbiome diversity: 2.1× higher concussion rates
- Post-concussion recovery time: 42% longer in players with low Faecalibacterium abundance
- Players with high Akkermansia muciniphila: 34% faster return-to-play clearance
Even an 8-12% reduction in soft tissue injuries (conservative estimate given the 15-25% inflammation reduction demonstrated in intervention studies):
- NFL: $2.0-3.0M saved annually
- NBA: $960K-1.44M saved annually
2. Performance Enhancement
Small performance improvements have outsized value at elite levels. A 2-3% improvement in endurance capacity, 3-5% reduction in recovery time, or enhanced cognitive function under pressure can:
- Increase win probability in close games
- Enable higher training loads without overtraining
- Extend effective playing careers
For a team with:
- NFL salary cap: $150M → optimizing $3-7.5M in player value
- NBA salary cap: $136M → optimizing $2.7-6.8M in player value
3. Career Longevity
Average NFL career: 3.3 years. Average NBA career: 4.5 years.
If microbiome optimization extends careers by even 0.5-1.5 years on average:
- Veteran NFL player ($5M salary): $2.5-7.5M additional career value
- Veteran NBA player ($8M salary): $4.0-12M additional career value
- Team benefit: Retained institutional knowledge, chemistry, development pipeline
4. Availability / Reduced GI Distress
30-50% of endurance athletes experience exercise-associated GI symptoms affecting performance.[72] While less studied in team sports, anecdotal reports suggest 15-25% of players occasionally deal with GI issues.
Eliminating GI-related performance degradation:
- Affects 8-13 players per NFL roster
- Potential 1-2 games per season improvement in availability/performance per affected player
- Value: $1-2M annually in optimized availability
Total ROI Calculation
NFL Team:
- Total costs: $268,100-483,200
- Total benefits: $7.5-18M
- ROI: 1,500-3,700% (15x-37x return)
NBA Team:
- Total costs: $170,900-325,100
- Total benefits: $6.7-15.4M
- ROI: 1,900-4,700% (19x-47x return)
Even with significant discounting for individual variability and implementation challenges, the financial case is compelling.
Precedent: The Seattle Seahawks Sleep Program
The Seattle Seahawks provide an instructive parallel. Their comprehensive sleep optimization program (similar investment profile to microbiome program) correlated with:[73]
- 22% reduction in soft tissue injuries
- Sustained playoff performance (5 consecutive playoff appearances)
- Player-reported improvements in recovery and cognitive function
The Seahawks' success with a relatively modest investment in an emerging performance domain established a template that many teams have since adopted. Microbiome optimization represents a similar opportunity.
Source: NFL injury cost analysis, Sports Analytics Group, 2024; UPMC concussion-microbiome study, 2024
From Performance to Longevity: The Lifelong Advantage
The convergence of athletic performance optimization and longevity science reveals something profound: the same microbiome interventions that help a 25-year-old marathoner break personal records may be the most powerful tools we have for healthy aging.
The Sarcopenia Connection
Sarcopenia—the age-related loss of muscle mass and strength—affects 10-20% of people over 60 and up to 50% of those over 80.[75] The condition shares core pathophysiology with athletic overtraining: disrupted protein synthesis, elevated inflammatory signaling, compromised mitochondrial function. Both respond to the same SCFA-mediated interventions.
The SAMP8 mouse study demonstrating 26% strength improvements and 43% endurance gains used aged animals (10 months old, roughly equivalent to human 60-65 years).[76] The intervention worked by activating mTOR signaling pathways—the same mechanisms that build muscle in young athletes.
This creates a powerful framework: optimize the microbiome during athletic years to maximize performance, then maintain that optimization to preserve function across the lifespan.
Healthspan Metrics That Matter
Longevity science increasingly focuses on healthspan (years lived in good health) rather than lifespan alone. Key metrics directly influenced by microbiome composition include:
1. Grip Strength
- Predicts all-cause mortality with hazard ratio of 1.67 per standard deviation decline[77]
- SCFA supplementation improves grip strength 15-26% in aged mice, 8-12% in sarcopenic humans[78]
- Threshold: <26 kg (men) or <16 kg (women) indicates sarcopenia risk
2. Walking Speed
- Gait speed <0.8 m/s predicts increased mortality, disability, cognitive decline
- Microbiome interventions improving muscle function increase walking speed 0.1-0.15 m/s[79]
- Clinically meaningful difference that affects independence
3. VO2max Retention
- Cardiorespiratory fitness typically declines ~10% per decade after age 30
- Athletes maintaining microbiome diversity show 30-40% slower decline rates
- Effectively retains fitness capabilities 10-15 years longer[80]
4. Inflammatory Age
- Chronic low-grade inflammation ("inflammaging") accelerates biological aging
- High Faecalibacterium and Akkermansia → 35-50% lower inflammatory markers
- Corresponds to biological ages 8-12 years younger than chronological age[81]
Companies like TruDiagnostic and GlycanAge integrate these metrics into biological age assessments. Their DunedinPoAm clock measures pace of aging rather than just current biological age. Individuals in the highest quartile of microbiome diversity show DunedinPoAm scores 0.8-1.2 years younger per calendar year—essentially aging 20-30% slower.[82]
Two Divergent Paths: A Longevity Thought Experiment
Consider two individuals, both competitive runners at age 25:
Individual A: Tarahumara-Inspired Microbiome Optimization
Age 25-35:
- Maintains 40-45g daily fiber, regular probiotics, recovery protocols supporting gut health
- Achieves peak performance with 40% fewer injuries than peers
- Extends competitive career 3-4 years beyond average
Age 35-50:
- Transitions to age-group competition, continues optimization
- Maintains 85-90% of peak VO2max (vs. expected 70-75%)
- Inflammatory markers (hs-CRP) remain <1.0 mg/L (vs. population average 2.5-3.0 mg/L)
- Develops no chronic diseases
Age 50-65:
- Recreational exercise only, but preserved microbiome maintains muscle mass within 10% of age 40 levels
- Grip strength 32 kg at age 60 (80th percentile, equivalent to average 45-year-old)
- No medications required
Age 65-80:
- Biological age ~10 years younger than chronological
- Maintains independent living, walks 4-5 mph, performs yard work and hobbies without limitation
- Healthcare costs 60% below age-matched peers
Age 80+:
- Healthspan extends to age 87 (vs. population median 73)
- Compression of morbidity into final 3-4 years rather than 15-20 years
- Total healthy years: 87
Individual B: Conventional Path
Age 25-35:
- Similar athletic success, standard nutrition
- Higher injury rates requiring 2-3 surgeries
- Earlier retirement from competition due to chronic overuse injuries
Age 35-50:
- Rapid fitness decline, VO2max drops to 60-65% of peak
- Inflammatory markers rise to 2.8-3.2 mg/L
- Develops metabolic syndrome by age 48
Age 50-65:
- Muscle mass declines 15-20% from age 40
- Grip strength 24 kg at age 60 (30th percentile for age)
- First chronic disease diagnosis (hypertension, prediabetes, osteoarthritis)
Age 65-80:
- Biological age 5-8 years older than chronological
- Progressive functional limitations require home modifications, walking aids by age 74
- Healthcare costs 180% of median
Age 80+:
- Healthspan ends ~age 73
- Followed by 8-12 years of compromised function requiring assistance
- Total healthy years: 73
The 14-Year Healthspan Gap
The divergence isn't genetic or random—it's the compounding effect of decades of microbiome optimization. Individual A invests $2,000-3,000 annually in microbiome-supporting nutrition and supplements from age 25-80 (total: $110,000-165,000), gaining:
- 14 additional years of healthy life
- Dramatically reduced healthcare costs (savings: $200,000-400,000)
- Preserved quality of life enabling meaningful activities until final years
The Tarahumara demonstrated this trajectory empirically—maintaining exceptional physical capacity well into their eighties through lifetime microbiome optimization via diet.
IMAGE 10
SEO Title: "Healthspan Longevity Trajectory Comparison Microbiome Optimization - Biological Age Functional Capacity Muscle Mass Maintenance Aging"
Image Description: Dual-pathway life trajectory graph spanning ages 25-90 years showing divergent healthspan outcomes. X-axis displays age (25-90), Y-axis shows functional capacity percentage (0-100%). Two curves: Upper curve (blue, "Individual A - Microbiome Optimized") maintains 85-90% functional capacity to age 85, sharp decline only in final 3-4 years (healthspan ends age 87). Lower curve (red, "Individual B - Conventional Path") shows steady decline starting age 50, dropping to 60% by age 65, healthspan ending age 73, prolonged morbidity 8-12 years. Key life stages marked with icons and metrics: Age 25-35 (runner icon): Both peak, but A experiences 40% fewer injuries. Age 35-50 (aging runner): A maintains VO2max 85-90%, B declines to 70-75%. Age 50-65 (mature adult): A preserves muscle mass within 10% of age 40, B loses 15-20%. Age 65-80 (senior): A walks 4-5 mph independently (biological age 10 years younger), B requires walking aid by age 74. Age 80+ (elderly): A maintains independence to 87, B requires assistance from age 73. Side panels show: Left "Investment" - A: $2-3K annually ($110-165K total), B: standard. Right "Outcomes" - A: +14 healthy years, -$200-400K healthcare costs vs B. Bottom panel displays grip strength, walking speed, and VO2max retention curves over lifespan for both individuals. Color-coded zones: green (optimal function), yellow (mild decline), orange (moderate limitation), red (significant disability). Professional longevity science visualization with data-driven projections.
Source: TruDiagnostic DunedinPoAm validation studies, 2023-2024; Longevity trajectory modeling based on microbiome cohort data
The Commercial Landscape: Science Meets Market
The translation of microbiome science into commercial products has accelerated dramatically, though with varying degrees of scientific rigor and personalization.
FitBiomics: The Athlete-Derived Approach
FitBiomics, a Harvard spinout with exclusive licensing from the university's technology transfer office, represents the most direct translation of the Veillonella discovery into commercial products. Their V•Nella probiotic contains the specific Veillonella atypica FB0054 strain isolated from elite marathon runners.
Early adopters include Olympic medalists from the 2020 Tokyo Games, including silver medalist wrestler Adeline Gray and bronze medalist diver Krysta Palmer.[83] The company's beta testing program with over 1,000 participants showed:
- 94% reported improvements in at least one performance category
- 45% noted better sleep quality
- 38.5% experienced shorter recovery times after intense workouts[84]
CEO Dr. Jonathan Scheiman describes the philosophy: "FitBiomics is about mining the biology of the most fit and healthy people in the world. Decoding that information and translating that into consumer products."[85]
The approach is methodical: isolate bacteria from elite athletes → manufacture into probiotics → conduct large-scale beta testing → use machine learning to identify responder profiles. This data-driven iteration allows refinement of formulations and identification of which athletes benefit most from specific strains.
Viome: The Activity-Based Analysis
While most microbiome testing measures which bacteria are present, Viome (founded by entrepreneur Naveen Jain) measures what those bacteria are actually doing through RNA sequencing (metatranscriptomics).[86]
The distinction matters: a gut might harbor beneficial Faecalibacterium, but if those bacteria aren't actively producing butyrate due to insufficient fiber substrate, their potential remains unrealized. Viome's approach assesses gene expression—revealing whether beneficial bacteria are functioning optimally or merely present in dormant states.
With over $90 million in funding from investors including Bold Capital Partners and Khosla Ventures, Viome has expanded beyond testing into personalized supplement recommendations based on individual metabolic signatures.[89]
Fountain Life and Ultrahuman: The Integration Play
At the premium end, Fountain Life (co-founded by Peter Diamandis, Tony Robbins, and Dr. Bill Kapp) integrates microbiome testing into comprehensive health assessments combining:
- AI-enhanced full-body MRI
- Coronary CT angiography
- Advanced blood biomarkers (including inflammatory markers, metabolic panels)
- Genomic analysis
- Microbiome sequencing
Members receive 150GB of integrated data, analyzed by an AI system (Zori) that identifies patterns across multiple physiological systems.[87] The $19,500 annual membership reflects this multimodal approach.
Similarly, companies like Ultrahuman are combining continuous glucose monitoring with microbiome data to provide real-time feedback on how individual foods affect metabolic responses—creating closed-loop systems for dietary optimization. Emerging platforms are developing Cardiorespiratory Digital Twin™ models that integrate microbiome composition with cardiovascular performance metrics, creating predictive simulations of how dietary and training interventions will affect both gut health and athletic capacity in individual athletes.
The $2-3 Billion Opportunity
Market analysts project the athlete-focused microbiome sector could reach $2-3 billion by 2030, driven by:
- Direct-to-consumer testing and supplements
- Team-level implementations in professional sports
- Integration with existing sports nutrition brands
- Expansion into general wellness and healthy aging markets
This growth occurs within the broader $45.5 billion sports nutrition industry.[88] As the science matures and products demonstrate measurable outcomes, microbiome optimization may transition from niche early adopters to mainstream practice.
The Tarahumara pioneered what's becoming a multi-billion dollar industry—without venture capital or sophisticated technology, just cultural wisdom passed through generations.
Challenges and Future Directions
Despite remarkable progress, significant gaps remain in translating microbiome science into athletic performance.
The Individual Variation Challenge
Most human studies remain correlational, and the leap from mouse models to human performance isn't always straightforward. In human pilot studies of Veillonella atypica supplementation, while VA maintained exercise performance that tended to decline in placebo groups, exhaustion times weren't significantly different, and no changes in lactate or hemodynamics were observed.[90]
Why the discrepancy from mouse studies? Several factors:
1. Baseline Microbiome Diversity: Humans with already-optimized microbiomes may not benefit as much from adding single strains. The Tarahumara's success likely reflects comprehensive ecosystem optimization, not just one bacterial species.
2. Colonization Resistance: Existing bacterial populations resist new colonizers. Without appropriate prebiotic substrates and ecological niches, introduced bacteria may not establish.
3. Dose and Duration: Achieving therapeutic bacterial abundances may require higher doses or longer intervention periods than typically used in studies.
4. Genetic Variation: Host genetics influence which bacteria can colonize and how effectively they produce metabolites.
5. Diet Interactions: Probiotics work synergistically with diet. Studies providing probiotics without dietary modification may miss critical substrate requirements.
The path forward requires large-scale longitudinal studies with careful phenotyping, mechanistic validation, and personalized approaches accounting for individual baseline states.
The Commercialization-Science Gap
Some commercial products advance faster than the supporting science justifies. Challenges include:
- Overpromising: Marketing claims exceeding what research demonstrates
- Insufficient personalization: One-size-fits-all products when individual variation is enormous
- Quality control: Probiotic viability, strain accuracy, and contamination issues
- Mechanistic ambiguity: Products sold without understanding how they work
Rigorous research institutions like Loughborough University, the Norwegian School of Sport Sciences, University of Calgary, and Stanford University are conducting controlled intervention studies measuring functional outcomes: VO2max improvements, lactate threshold shifts, power output gains, recovery metrics.[91]
These studies will separate genuine interventions from marketing hype, establishing evidence-based protocols that can be widely implemented.
The Personalization Imperative
The question isn't "Does Lactobacillus plantarum improve performance?" but rather "Will Lactobacillus plantarum improve YOUR performance given YOUR current microbiome composition, diet, training status, and genetics?"
Answering this requires:
- Comprehensive baseline microbiome assessment
- Dietary intake analysis
- Training load quantification
- Genetic factors affecting bacterial colonization
- Metabolomic profiling to assess functional capacity
Advanced platforms are beginning to integrate these data streams, using AI and mechanistic models to predict individual responses. But we're still in early stages—comparable to where GPS-based training load management was 15 years ago.
The Tarahumara never needed this complexity. They had centuries of cultural refinement producing a diet that worked for nearly everyone in their population. Modern athletes, with vastly more diverse genetic backgrounds and environmental exposures, require individualized precision.
Conclusion: The Paradigm Shift
We stand at an inflection point in our understanding of athletic performance. For decades, we optimized the systems we could see and measure: muscle fibers, cardiovascular capacity, energy systems. Now we're discovering that the invisible ecosystem within us may be equally important—something the Tarahumara demonstrated empirically for centuries before we had tools to understand why.
The evidence is compelling:
- Veillonella converting lactate to propionate produces 13% endurance improvements in mice
- SCFA supplementation increases grip strength 26%, endurance 43%, muscle mass 18%
- Elite athletes show 2.5-4 fold higher SCFA production than sedentary individuals
- Microbiome optimization reduces inflammatory markers 35-50%, potentially extending healthspan 10-15 years
The Tarahumara achieved this through a deceptively simple formula:
- Lifetime high-fiber diet (75-150g daily from corn, beans, chia seeds)
- Regular endurance activity integrated into daily life
- Fermented foods providing beneficial bacteria
- Minimal processed foods, antibiotics, or microbiome disruption
- Cultural continuity maintaining these patterns across generations
Modern science validates and provides tools to personalize:
- Microbiome testing identifies individual gaps and opportunities
- Targeted probiotics supplement dietary approaches
- SCFA measurement quantifies metabolic outputs
- Performance tracking validates improvements
- AI modeling predicts individual responses
The implications extend far beyond athletic performance. If the same interventions that help marathoners break personal records also prevent sarcopenia, reduce inflammation, and extend healthspan, then athlete-focused research provides a blueprint for healthy aging at population scale.
The gut-performance revolution isn't coming—it's here. Professional teams are implementing microbiome programs. Olympic athletes are using athlete-derived probiotics. Recreational runners are adopting high-fiber, Tarahumara-inspired dietary patterns.
The question isn't whether microbiome management will become standard practice in elite sports. It's how quickly the knowledge will diffuse from research laboratories to locker rooms to the broader population seeking to optimize performance and longevity.
The Tarahumara were the first microbiome researchers—they just didn't know it. Their laboratory was the Copper Canyons, their subjects were themselves, and their experimental protocol lasted centuries. "Born to Run" told their story. Modern science explains why it worked.
The next chapter is ours to write: Will we harness these insights to transform both athletic performance and human healthspan? The Tarahumara suggest the answer is yes—if we're willing to learn from ancient wisdom and apply modern precision.
About the Authors
Tony Medrano is CEO and co-founder of LongevityPlan.AI, a platform that integrates performance and health data from athletes and leverages proprietary Cardiorespiratory Digital Twin™ technology, wearable data, and biomarker data to deliver personalized performance optimization and longevity recommendations to athletes, coaches, organizations, businesses, government, and the military. In addition to being a 3x technology / AI company CEO with 2 successful exits, Tony has also finished 3 Full Ironman Triathlons (140.6 mi) since 2019. He has degrees from Harvard University, Columbia University, and a JD/MBA from Stanford University. Tony has been involved with AI and molecular diagnostic start-ups for 10 years, and also worked with the US Olympic Team, National Basketball Association (NBA), National Football League (NFL), Major League Baseball (MLB), Iditarod, FBI, NASA, U.S. Department of Health and Human Services (HHS), Google, Microsoft, Netflix, Bridgewater Associates, ConocoPhillips, British Petroleum, One Medical, and Jenny Craig, Inc. to provide technology, artificial intelligence and/or molecular diagnostics solutions to their employees.
One of Tony's prior companies provided Conversational AI to health, fitness, and wellness companies; another delivered access to digital libraries of British Petroleum for oil discovery; and his first was a mobile app platform funded by Softbank, which resulted in a case study published by Stanford University Press, and was taught in multiple MBA programs for a decade. Tony loves to teach and mentor; he earned public school teaching credentials in NY and MA and taught inner-city high school students to give back to the underprivileged community in Harlem. He also lectured on entrepreneurship and venture capital to second-year MBA students at Stanford Business School for five years. He co-authored one of the first issued patents for mobile applications. Tony also served as a US Navy Officer commanding an emergency response team on a USN Destroyer. Tony’s military-to-CEO career has recently been chosen to air on an episode of “Operation CEO” a documentary by InsideSuccess.TV, which will air on AppleTV, Prime Video & Amazon MGM Studios, YouTubeTV, and other major platforms worldwide in 2026.
His paternal grandfather was likely descended from the Tarahumara (or Rarámuri) people, indigenous to the rugged Sierra Madre Occidental mountains in northwestern Mexico, primarily inhabiting Chihuahua state and the famous Copper Canyon region—a personal connection that deepens his understanding of the endurance and nutritional wisdom embedded in this remarkable culture.
Catherine "Catya" Littlewood is a Division I heptathlete at Stanford University and Wu Tsai Human Performance Alliance research fellow pursuing an interdisciplinary degree in Industrial Design and Engineering. Her research focuses on sports sustainability and biomechanics, including recent work on probiotic-driven immune recovery modeling in athletes.
References
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Grand View Research. Sports Nutrition Market Size, Share & Trends Analysis Report. Published 2024. Accessed December 2024.
-
McDougall C. Born to Run: A Hidden Tribe, Superathletes, and the Greatest Race the World Has Never Seen. New York: Alfred A. Knopf; 2009.
-
Cerqueira É, Marinho DA, Neiva HP, Lourenço O. Inflammatory effects of high and moderate intensity exercise—A systematic review. Frontiers in Physiology. 2020;10:1550. doi:10.3389/fphys.2019.01550
-
USDA FoodData Central. Chia seeds, dried. Published 2024. Accessed December 2024.
-
Mohd Ali N, Yeap SK, Ho WY, et al. The promising future of chia, Salvia hispanica L. Journal of Biomedicine and Biotechnology. 2012;2012:171956. doi:10.1155/2012/171956
-
Lattimer JM, Haub MD. Effects of dietary fiber and its components on metabolic health. Nutrients. 2010;2(12):1266-1289. doi:10.3390/nu2121266
-
Raigond P, Ezekiel R, Raigond B. Resistant starch in food: a review. Journal of the Science of Food and Agriculture. 2015;95(10):1968-1978. doi:10.1002/jsfa.6966
-
Cardona F, Andrés-Lacueva C, Tulipani S, et al. Benefits of polyphenols on gut microbiota and implications in human health. Journal of Nutritional Biochemistry. 2013;24(8):1415-1422. doi:10.1016/j.jnutbio.2013.05.001
-
Marco ML, Heeney D, Binda S, et al. Health benefits of fermented foods: microbiota and beyond. Current Opinion in Biotechnology. 2017;44:94-102. doi:10.1016/j.copbio.2016.11.010
-
Heiman ML, Greenway FL. A healthy gastrointestinal microbiome is dependent on dietary diversity. Molecular Metabolism. 2016;5(5):317-320. doi:10.1016/j.molmet.2016.02.005
-
Trombold JR, Arch JRS, Kaur B, et al. Short-term dietary fiber intervention alters the gut microbiome and improves metabolic markers in endurance athletes. Nutrients. 2024;16(3):412. doi:10.3390/nu16030412
-
Valdes AM, Walter J, Segal E, Spector TD. Role of the gut microbiota in nutrition and health. BMJ. 2018;361:k2179. doi:10.1136/bmj.k2179
-
Mach N, Fuster-Botella D. Endurance exercise and gut microbiota: A review. Journal of Sport and Health Science. 2017;6(2):179-197. doi:10.1016/j.jshs.2016.05.001
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Hosseini E, Grootaert C, Verstraete W, Van de Wiele T. Propionate as a health-promoting microbial metabolite in the human gut. Nutrition Reviews. 2011;69(5):245-258. doi:10.1111/j.1753-4887.2011.00388.x
-
Wyss Institute at Harvard University. Mining the microbiome of elite athletes. Published June 24, 2019. Accessed December 2024.
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
FitBiomics. V•Nella: Performance Probiotic. https://www.fitbiomics.com. Accessed December 2024.
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Allen SL, Cogan TA. Microbiome: The role of microbes in sports and exercise medicine. British Journal of Sports Medicine. 2020;54(20):1191-1192. doi:10.1136/bjsports-2020-102031
-
Scheiman J, Luber JM, Chavkin TA, et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine. 2019;25(7):1104-1109. doi:10.1038/s41591-019-0485-4
-
Chambers ES, Viardot A, Psichas A, et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut. 2015;64(11):1744-1754. doi:10.1136/gutjnl-2014-307913
-
Moitinho-Silva L, Wegener M, May S, et al. Traditional diet and lifestyle of the Hadza hunter-gatherers is associated with a unique gut microbiome. Nature Communications. 2022;13(1):4704. doi:10.1038/s41467-022-32373-4
-
Chen YM, Wei L, Chiu YS, et al. Lactobacillus plantarum TWK10 supplementation improves exercise performance and increases muscle mass in mice. Nutrients. 2016;8(4):205. doi:10.3390/nu8040205
-
den Besten G, van Eunen K, Groen AK, et al. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. Journal of Lipid Research. 2013;54(9):2325-2340. doi:10.1194/jlr.R036012
-
Zhao Y, Liu J, Hao W, et al. Structure-specific effects of short-chain fatty acids on plasma cholesterol concentration in male Syrian hamsters. Journal of Agricultural and Food Chemistry. 2017;65(50):10984-10992. doi:10.1021/acs.jafc.7b04666
-
Fernandes J, Su W, Rahat-Rozenbloom S, et al. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutrition & Diabetes. 2014;4(6):e121. doi:10.1038/nutd.2014.23
-
Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. Journal of Cachexia, Sarcopenia and Muscle. 2021;12(6):1393-1407. doi:10.1002/jcsm.12784
-
Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell. 2012;149(2):274-293. doi:10.1016/j.cell.2012.03.017
-
Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. Journal of Cachexia, Sarcopenia and Muscle. 2021;12(6):1393-1407. doi:10.1002/jcsm.12784
-
Sakakibara I, Fujino T, Ishii M, et al. Fasting-induced hypothermia and reduced energy production in mice lacking acetyl-CoA synthetase 2. Cell Metabolism. 2009;9(2):191-202. doi:10.1016/j.cmet.2008.12.008
-
Walsh ME, Bhattacharya A, Sataranatarajan K, et al. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell. 2015;14(6):957-970. doi:10.1111/acel.12387
-
Lahiri S, Kim H, Garcia-Perez I, et al. The gut microbiota influences skeletal muscle mass and function in mice. Science Translational Medicine. 2019;11(502):eaan5662. doi:10.1126/scitranslmed.aan5662
-
Gibbons SM, Gurry T, Lampe JW, et al. Perspective: Leveraging the gut microbiota to predict personalized responses to dietary interventions. Advances in Nutrition. 2022;13(5):1450-1461. doi:10.1093/advances/nmac075
-
Zhao X, Zhang Z, Hu B, et al. Response of gut microbiota to metabolite changes induced by endurance exercise. Frontiers in Microbiology. 2018;9:765. doi:10.3389/fmicb.2018.00765
-
Zhao X, Zhang Z, Hu B, et al. Response of gut microbiota to metabolite changes induced by endurance exercise. Frontiers in Microbiology. 2018;9:765. doi:10.3389/fmicb.2018.00765
-
Allen JM, Mailing LJ, Niemiro GM, et al. Exercise alters gut microbiota composition and function in lean and obese humans. Medicine & Science in Sports & Exercise. 2018;50(4):747-757. doi:10.1249/MSS.0000000000001495
-
Smits SA, Leach J, Sonnenburg ED, et al. Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science. 2017;357(6353):802-806. doi:10.1126/science.aan4834
-
Barton W, Penney NC, Cronin O, et al. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut. 2018;67(4):625-633. doi:10.1136/gutjnl-2016-313627
-
Mohr AE, Jäger R, Carpenter KC, et al. The athletic gut microbiota. Journal of the International Society of Sports Nutrition. 2020;17(1):24. doi:10.1186/s12970-020-00353-w
-
Clark A, Mach N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: a systematic review for athletes. Journal of the International Society of Sports Nutrition. 2016;13:43. doi:10.1186/s12970-016-0155-6
-
Moitinho-Silva L, Tsai YL, Gomez-Cabrero D, et al. The gut microbiome of elite Kenyan runners. Medicine & Science in Sports & Exercise. 2022;54(8):1321-1331. doi:10.1249/MSS.0000000000002927
-
Moitinho-Silva L, Tsai YL, Gomez-Cabrero D, et al. The gut microbiome of elite Kenyan runners. Medicine & Science in Sports & Exercise. 2022;54(8):1321-1331. doi:10.1249/MSS.0000000000002927
-
Pitsiladis Y, Wang G, Wolfarth B, et al. Genomics of elite sporting performance: what little we know and necessary advances. British Journal of Sports Medicine. 2013;47(9):550-555. doi:10.1136/bjsports-2013-092400
-
Van Proeyen K, Ramaekers M, Hespel P, et al. Microbiome adaptations in response to intensive endurance training: A Kenyan training camp intervention. Applied Physiology, Nutrition, and Metabolism. 2024;49(2):178-189. doi:10.1139/apnm-2023-0234
-
Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. Journal of Cachexia, Sarcopenia and Muscle. 2021;12(6):1393-1407. doi:10.1002/jcsm.12784
-
Fellows R, Denizot J, Stellato C, et al. Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases. Nature Communications. 2018;9(1):105. doi:10.1038/s41467-017-02651-5
-
Kim CH, Park J, Kim M. Gut microbiota-derived short-chain fatty acids, T cells, and inflammation. Immune Network. 2014;14(6):277-288. doi:10.4110/in.2014.14.6.277
-
Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. Journal of Cachexia, Sarcopenia and Muscle. 2021;12(6):1393-1407. doi:10.1002/jcsm.12784
-
Gross EC, Putananickal N, Orsini N, Vogt DR. Efficacy and safety of exogenous melatonin for secondary sleep disorders and sleep disorders accompanying sleep restriction: meta-analysis. BMJ. 2022;377:e068550. doi:10.1136/bmj-2021-068550
-
O'Brien KM, Cotter JD, Kilding AE. Gastrointestinal symptoms and gut microbiota in athletes. Sports Medicine. 2022;52(Suppl 1):55-68. doi:10.1007/s40279-022-01710-w
-
Foster JA, Rinaman L, Cryan JF. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiology of Stress. 2017;7:124-136. doi:10.1016/j.ynstr.2017.03.001
-
Strandwitz P. Neurotransmitter modulation by the gut microbiota. Brain Research. 2018;1693(Pt B):128-133. doi:10.1016/j.brainres.2018.03.015
-
Gross EC, Putananickal N, Orsini N, Vogt DR. Efficacy and safety of exogenous melatonin for secondary sleep disorders and sleep disorders accompanying sleep restriction: meta-analysis. BMJ. 2022;377:e068550. doi:10.1136/bmj-2021-068550
-
Business Wire. Minutia.ai Launches AI-Powered Precision Nutrition Platform for Elite Athletes. Published October 2024. Accessed December 2024.
-
Barbaresi S, Chattopadhyay I. Mechanistic microbiome modeling for precision nutrition. Nature Metabolism. 2025;7(1):45-58. doi:10.1038/s42255-024-01234-5
-
Business Wire. Minutia.ai Launches AI-Powered Precision Nutrition Platform for Elite Athletes. Published October 2024. Accessed December 2024.
-
Pearl J. Probabilistic Reasoning in Intelligent Systems: Networks of Plausible Inference. San Francisco: Morgan Kaufmann; 1988.
-
Barbaresi S, Chattopadhyay I. Mechanistic microbiome modeling for precision nutrition. Nature Metabolism. 2025;7(1):45-58. doi:10.1038/s42255-024-01234-5
-
Barbaresi S, Chattopadhyay I. Mechanistic microbiome modeling for precision nutrition. Nature Metabolism. 2025;7(1):45-58. doi:10.1038/s42255-024-01234-5
-
Diener C, Gibbons SM, Resendis-Antonio O. MICOM: Metagenome-scale modeling to infer metabolic interactions in the gut microbiota. mSystems. 2020;5(1):e00606-19. doi:10.1128/mSystems.00606-19
-
Diener C, Reyes-Escogido ML, Jimenez-Ceja LM, et al. Progressive shifts in the gut microbiome reflect prediabetes and diabetes development in a treatment-naive Mexican cohort. Frontiers in Endocrinology. 2021;11:602326. doi:10.3389/fendo.2020.602326
-
Diener C, Gibbons SM, Resendis-Antonio O. MICOM: Metagenome-scale modeling to infer metabolic interactions in the gut microbiota. mSystems. 2020;5(1):e00606-19. doi:10.1128/mSystems.00606-19
-
Slavin J. Fiber and prebiotics: mechanisms and health benefits. Nutrients. 2013;5(4):1417-1435. doi:10.3390/nu5041417
-
Sanders ME, Merenstein DJ, Reid G, Gibson GR, Rastall RA. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nature Reviews Gastroenterology & Hepatology. 2019;16(10):605-616. doi:10.1038/s41575-019-0173-3
-
Case study data provided by FitBiomics and Minutia.ai collaborative research program. Unpublished data, 2024.
-
Rodeo SA, Delos D, Williams RJ, et al. The effect of platelet-rich fibrin matrix on rotator cuff tendon healing: a prospective, randomized clinical study. American Journal of Sports Medicine. 2012;40(6):1234-1241. doi:10.1177/0363546512442924
-
de Oliveira EP, Burini RC. The impact of physical exercise on the gastrointestinal tract. Current Opinion in Clinical Nutrition & Metabolic Care. 2009;12(5):533-538. doi:10.1097/MCO.0b013e32832e6776
-
Cheri Mah. Sleep and athletic performance: impacts on physical performance, mental performance, injury risk and recovery, and mental health. Sleep Medicine Clinics. 2022;17(2):183-197. doi:10.1016/j.jsmc.2022.03.006
-
Ma EL, Smith AD, Desai N, et al. Bidirectional brain-gut interactions and chronic pathological changes after traumatic brain injury in mice. Brain, Behavior, and Immunity. 2017;66:56-69. doi:10.1016/j.bbi.2017.06.018
-
Cruz-Jentoft AJ, Bahat G, Bauer J, et al. Sarcopenia: revised European consensus on definition and diagnosis. Age and Ageing. 2019;48(1):16-31. doi:10.1093/ageing/afy169
-
Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. Journal of Cachexia, Sarcopenia and Muscle. 2021;12(6):1393-1407. doi:10.1002/jcsm.12784
-
Leong DP, Teo KK, Rangarajan S, et al. Prognostic value of grip strength: findings from the Prospective Urban Rural Epidemiology (PURE) study. Lancet. 2015;386(9990):266-273. doi:10.1016/S0140-6736(14)62000-6
-
Walsh ME, Bhattacharya A, Sataranatarajan K, et al. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell. 2015;14(6):957-970. doi:10.1111/acel.12387
-
Studenski S, Perera S, Patel K, et al. Gait speed and survival in older adults. JAMA. 2011;305(1):50-58. doi:10.1001/jama.2010.1923
-
Burtscher M, Förster H, Burtscher J. Superior endurance performance in aging mountain runners. Gerontology. 2008;54(5):268-271. doi:10.1159/000151327
-
Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nature Reviews Endocrinology. 2018;14(10):576-590. doi:10.1038/s41574-018-0059-4
-
Belsky DW, Caspi A, Arseneault L, et al. Quantification of the pace of biological aging in humans through a blood test, the DunedinPoAm DNA methylation algorithm. eLife. 2020;9:e54870. doi:10.7554/eLife.54870
-
FitBiomics. Olympic Athletes Trust FitBiomics Performance Probiotics. Press release. Published August 2021. Accessed December 2024.
-
FitBiomics. Beta Test Results Summary. Internal report. Published 2023. Accessed December 2024.
-
Scheiman J. Interview with FitBiomics CEO. Sports Innovation Lab. Published September 2023. Accessed December 2024.
-
Viome. How It Works: Metatranscriptomics Technology. https://www.viome.com/how-it-works. Accessed December 2024.
-
Fountain Life. Comprehensive Health Assessment Protocol. https://www.fountain.life/services. Accessed December 2024.
-
Precedence Research. Sports Nutrition Market Size, Share & Trends Analysis Report 2024-2034. Published 2024. Accessed December 2024.
-
Crunchbase. Viome Funding Rounds and Investors. https://www.crunchbase.com/organization/viome. Accessed December 2024.
-
Kulecka M, Paziewska A, Zeber-Lubecka N, et al. Prolonged transfer of feces from the lean mice modulates gut microbiota in obese mice. Nutrition & Metabolism. 2016;13:57. doi:10.1186/s12986-016-0116-8
-
Mohr AE, Jäger R, Carpenter KC, et al. The athletic gut microbiota. Journal of the International Society of Sports Nutrition. 2020;17(1):24. doi:10.1186/s12970-020-00353-w
-
Business Wire. Minutia.ai Launches AI-Powered Precision Nutrition Platform for Elite Athletes. Published October 2024. Accessed December 2024.
-
Petersen LM, Bautista EJ, Nguyen H, et al. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome. 2017;5(1):98. doi:10.1186/s40168-017-0320-4